




















 [6]
[6]
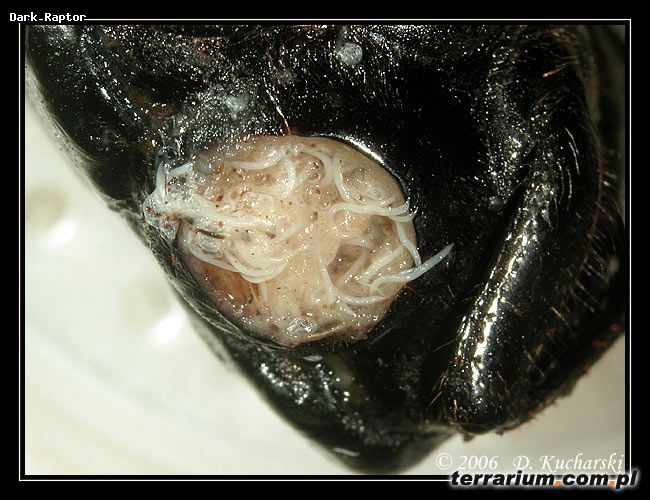
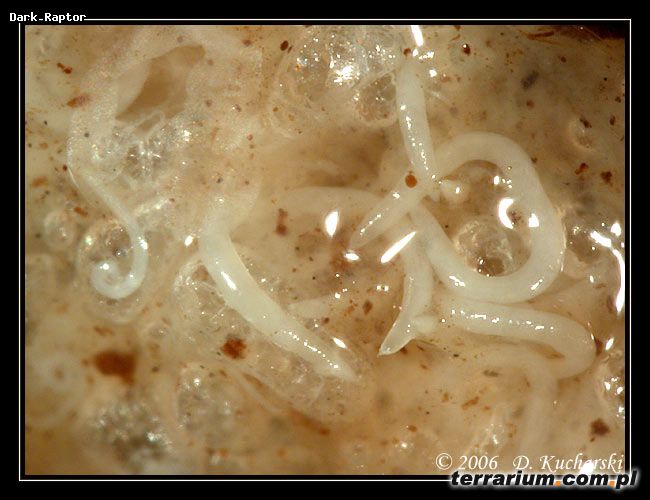


Nicienie entomopatogenne
Naturalne infekcje owadów przez entomopatogenne nicienie znane są już od kilkudziesięciu lat. Opisano 23 rodziny nicieni pasożytujących na owadach (min. Mermithidae, Tetradonematidae (rząd: Mermithidae); Allantonematidae, Phaenopsitylenchidae i Sphaeruliidae (rząd: Tylenchida); Heterorhabditidae i Steinernematidae (rząd: Rhabditida)).
Po zainfekowaniu owada nicienie odżywiają się głównie bakteriami namnażającymi się w jego tkankach. Cykl rozwojowy pasożytów trwa od 7-10 dni (Steinernematidae) do 15 dni (Heterorhabditidae). Po zakończeniu cyklu (jednego lub kilku) oraz wyczerpaniu zapasów pokarmowych larwy inwazyjne opuszczają ciało owada – żywiciela. Migracja larw zostaje zahamowana podczas niekorzystnych warunków środowiskowych (niska temperatura i wilgotność gleby). Okres wstrzymania wędrówki może wynosić nawet 50 dni.
Po opuszczeniu martwego już żywiciela larwy nicieni aktywnie poszukują następnego. Istnieje szereg czynników pomagających pasożytom odnaleźć owada. Należą do nich min: wzrastające stężenie CO2 i temperatura, obecność odchodów. Drogi którymi nicienie wnikają do wnętrza ofiary mogą być różne: przez przetchlinki, otwór pokarmowy i odbytowy oraz przez miękki oskórek okrywający ciało. Owady w toku ewolucji rozwinęły mechanizmy utrudniające ich odnalezienie przez patogeny. Należą do nich min: wysoki współczynnik defekacji (niskie ryzyko zarażenia przez odbyt) u larw żukowatych; budowanie ścian oddzielających zarażone osobniki od zdrowych współmieszkańców u termitów.
Głównymi miejscami docelowymi pasożytów w organizmie owada są: jelito, jama ciała, układ rozrodczy i cewki Malpighiego.
Życie w środowisku, w którym pasożyty stanowią dużą część fauny wpłynęło na pojawienie się u owadów szeregu mechanizmów odpornościowych:
1. odporności mechanicznej przeciwinwazyjnej (bariery anatomiczno – fizjologiczne)
2. odporności komórkowej (fagocytoza, obronne odczyny hemocytarne: nodulacje, inkapsulacja)
3. odporności humoralnej (bakteriobójcze czynniki hemolimfy)
1. Bariery anatomiczno – fizjologiczne
Ten rodzaj odporności ukierunkowany jest na niedopuszczenie entomopatogenów do jamy ciała.
Ciało owada osłonięte jest przez szkielet zewnętrzny zbudowany z chityny i białek. Stanowi on twardą, chemiczną barierę ochronną dla wielu pasożytów. Jelito przednie i tylne posiada wyściółkę chitynową, która chroni przez wniknięciem zarazków do jamy ciała. Z kolei jelito środkowe zaopatrzone jest w błonę perytroficzną oraz nabłonek pokryty warstwą śluzu, które uniemożliwiają przemieszczanie się wirusów, bakterii i pasożytów. Działanie chemiczne hamujące rozwój patogenów mają: pH treści jelitowej, substancje antybiotyczne oraz enzymy bakteriolityczne wytwarzane przez symbiotyczną mikroflorę jelitową owada. Otwór gębowy zaopatrzony jest w chitynowe filtry hamujące wnikanie pasożytów do środka. Układ tchawkowy oprócz obecności chitynowych sit charakteryzuje się niską wilgotnością i brakiem substancji pokarmowych, co stwarza niekorzystne warunki do kolonizacji i rozwoju patogenów. Przełamanie tych barier następuje tylko przy współpracy nicieni i bakterii. Wspólnie produkują one enzymy chitynolityczne i dezaktywujące białka owadzie, co tworzy wrota zakażenia. Uszkodzenie pierwszej linii obrony aktywuje mechanizmy obrony wewnętrznej owada. Po rozpoznaniu ciała obcego organizm ofiary uruchamia natychmiast reakcję odpornościową. Następuje interakcja czynników komórkowych i humoralnych.
2. Odporność komórkowa
Odporność wewnętrzna chroni organizm owada przed zakażeniem saprofitami i jest mniej skuteczna w stosunku do drobnoustrojów owadobójczych.
Po rozpoznaniu ciał obcych, organizm uruchamia kaskadę reakcji obronnych w celu przywrócenia równowagi ustroju.
a) Fagocytoza
Mechanizm fagocytozy składa się z kilku etapów: rozpoznanie substancji jako obcej dla organizmu, chemotaksja – ruch w kierunku patogenu, pochłanianie (endocytoza), wewnętrzne procesy metaboliczne. Celem fagocytozy jest strawienie obcej substancji przez specyficzne komórki owada (hemocyty – plazmocyty i granulocyty) lub jej odłożenie w organellach komórkowych pełniących funkcję „zbiorników odpadów”. Fagocytoza jest skutecznym mechanizmem obronnym dopóki nie zostanie przekroczone stężenie progowe zarazków, indywidualne dla każdego gatunku.
b) Nodulacja
Po przekroczeniu powyższej wartości progowej fagocytoza jest wspomagana przez nodulację (tworzenie guzków). Komórki fagocytarne wypełnione obcym materiałem są otaczane kilkoma warstwami hemocytów, a następnie ulegają melanizacji (synteza barwnika – melaniny). Zmelanizowane guzki izolują obce substancje od płynów jamy ciała.
c) Inkapsulacja
Drobne cząsteczki o średnicy poniżej 10µm (wirusy, bakterie) są fagocytowane przez pojedyncze hemocyty. Cząsteczki o średnicy przekraczającej 10µm (pasożyty i ich jaja, strzępki grzybni) ulegają inkapsulacji. Substancja obca wraz z otoczką hemocytarną tworzy kapsułę. Końcowym efektem jest sekwestracja pasożytów, czyli ich wychwycenie, transport oraz unieszkodliwienie. Proces inkapsulacji jest inicjowany pod wpływem czynników zranienia (injury factors), uwalnianych przez uszkodzone i patologicznie zmienione komórki gospodarza.
Owady nie posiadają przeciwciał oraz pamięci immunologicznej jak kręgowce, ale ich reakcje komórkowe i humoralne pojawiają się szybko i działają krótko likwidując zakażenie. Taki system zapewniają owadom antysomy (indukowane czynniki przeciwbakteryjne hemolimfy) które aktywują reakcje odpornościowe.
3. Odporność humoralna
Drugim typem odporności wewnętrznej jest odporność humoralna. Uwarunkowana jest ona obecnością w hemolimfie czynników rozpuszczalnych niszczących patogeny.
Główną funkcję w tym typie odporności pełni lizozym. Synteza tego związku ma miejsce w ciele tłuszczowym skąd przechodzi do hemolimfy. Podwyższonemu poziomowi lizozymu w hemolimfie towarzyszy wzrost działania ochronnego organizmu. Lizozym współpracuje z białkami hemolimfy (cekropiny i attacyny). Główną funkcją lizozymu jest usuwanie mureiny, składnika ściany komórkowej bakterii (głównie gram – dodatnich), zabitych uprzednio przez cekropiny. Cekropiny niszczą również bakterie gram – ujemne. Cząsteczki cekropiny są syntetyzowane pod wpływem sygnału pochodzącego z hemocytów wypełnionych sfagocytowanymi bakteriami, które po kontakcie z komórkami ciała tłuszczowego pobudzają je do syntezy mRNA odpornościowego, który z kolei aktywuje szereg reakcji katalizujących powstanie cekropin. Lizozym i cekropiny oczyszczają krew owada. W usuwaniu zarazków istotną rolę odgrywają również attacyny, które osłabiają ścianę komórkową patogenów, czyniąc ją bardziej podatną na działanie cekropin i lizozymu.
Lizozym występuje również w szkielecie zewnętrznym. Istnieje zależność pomiędzy stężeniem lizozymu w oskórku a sposobem życia lub budową ciała owada. Owady z twardym szkieletem zewnętrznym zawierają niewielką ilość lizozymu, z kolei te z miękką okrywą ciała i bytujące w środowisku obfitym w bakterie posiadają wysokie stężenia lizozymu w oskórku (Lepidoptera, Diptera).
Pasożyty i patogeny owadów posiadają również szereg mechanizmów przełamywania odporności owadów, do których zaliczamy:
I. uszkadzanie struktur owada przez toksyny i enzymy histolityczne syntetyzowane przez patogena. Mogą one zahamować obronę przeciwzakaźną gospodarza i zwykle kończą się jego śmiercią.
II. nieobecności skutecznych mechanizmów kontroli immunologicznej patogena w ciele owada. Patogen może się rozwijać ponieważ nie jest niszczony w hemolimfie owada (brak działań obronnych).
III. mechanizmom ucieczki spod kontroli immunologicznej oraz niszczenie lub modyfikacja składowych odporności komórkowej i humoralnej.
Ucieczka spod kontroli immunologicznej możliwa jest dzięki oporowi biernemu lub czynnemu patogenów. Do oporu biernego zaliczymy np. mimikrę molekularną (upodobnienie struktur powierzchniowych patogena, będących markerami swoistości immunologicznej, do struktur owada), kolonizację regionów ciała owada o niskiej reaktywności, rozwój w narządach oddalonych od obszarów dużej koncentracji hemocytów. Zahamowanie reakcji komórkowej wymaga obecności jadu pasożyta. Zmiana fazy wzrostu bakterii również osłabia odporność owada. Dymorfizm morfologiczny polega na przejściu z formy gładkiej w formę szorstką, co utrudnia aktywność fagocytozy oraz działanie bakteriobójcze hemolimfy.
Niektóre metabolity pasożytów hamują inkapsulację w jamie ciała owada.
Opór czynny pozwala nicieniom entomopatogennym oraz innym patogenom na uniknięcie fagocytozy lub przetrwanie po sfagocytozowaniu. Takie działanie jest możliwe dzięki syntezie agresyn (substancje odstraszające fagocyty) lub przez zmianę struktury powierzchni komórki bakteryjnej, która staje się niewrażliwa na enzymy lizosomalne. Sfagocytowane patogeny są zdolne nawet do namnażania się wewnątrz fagocytów. Przełamanie barier odporności humoralnej owada możliwe jest dzięki egzoproteinazom bakteryjnym lub inhibitorom odporności typu proteaz wytwarzanym przez nicienie. Opór czynny to również inhibicja chemotaksji, fagocytozy, inkapsulacji oraz blokowanie aktywacji układu profenylooksydazy. Mechanizmy tych oddziaływań są jeszcze słabo poznane.
Zjadliwość (wirulencja) patogenów jest uwarunkowana szeregiem właściwości przełamywania systemów odpornościowych owadów, zdolnością szybkiego rozmnażania się oraz kolonizacji tkanek zarażonego organizmu.
Opracowała Kornelia Kucharska na podstawie literatury:
Jarosz J.1996. Strategie obrony przeciwzakaźnej i sposoby ucieczki entomopatogenów spod kontroli immunologicznej owadów. Wiadomości Parazytologiczne 42: 3-27
Jaworska M.1993. Wpływ owadobójczych nicieni z rodziny Heterorhabditidae i Steinernematidae na śmietkę kapuścianą Delia brassicae Hoffmannsegg (Dietera, Anthomyiidae) i jej wrogów naturalnych. Polskie Pismo Entomologiczne 62: 243-254.
Kowalska J. Wzajemne powiązania pomiędzy nicieniami owadobójczymi, owadami i bakteriami oraz ich wykorzystanie w praktyce. 93- 98
Liczba wyświetleń: 5876

















