




















 [6]
[6]
Pasożyty zwierzęce płazów
W artykule zaprezentowano wybrane grupy pasożytów
Pasożyty (helminofauna) stanowią ok. 25% wszystkich dotychczas opisanych gatunków zwierząt. Termin pasożytnictwo został po raz pierwszy wprowadzony do nauki w XIX w. Definicję sformułował Leuckart w 1852 r. Pojęcie odnosiło się tylko do organizmów wykorzystujących inne organizmy, bez ich zabijania, w celu zdobycia pokarmu. Powyższa definicja jest uproszczona i obecnie stale ulega modyfikacji, pojawiają się nowe jej aspekty. Ciekawostką jest to, że pasożytnictwo można uznać za specyficzną formę drapieżnictwa. Oba układy różnią się jednak zasadniczo. W relacji drapieżnik – ofiara liczba wydawanego na świat potomstwa drapieżnika jest niższa niż u jego ofiary. Ofiara zostaje najczęściej natychmiast uśmiercona i spożyta. Z kolei w zależności pasożyt – żywiciel liczba wydawanego na świat potomstwa pasożyta jest wyższa niż u jego gospodarza. Interakcja ta, ma najczęściej długotrwały charakter i polega na powolnej eksploatacji żywiciela.
W zależności od otoczenia, w którym pasożyt aktualnie przebywa wyróżniamy:
- mikrośrodowisko – ciało żywiciela
- makrośrodowisko – środowisko zewnętrzne, poza ciałem ofiary.
Z lokalizacją helmintofauny związane są następujące rodzaje pasożytnictwa:
- ektopasożytnictwo – pasożyty zewnętrzne bytują na powierzchni ciała żywiciela lub w jego jamach mających kontakt ze środowiskiem zewnętrznym, np. zewnętrzny przewód słuchowy, jama nosowa, skrzela.
- endopasożytnictwo – pasożyty wewnętrzne bytują wewnątrz ciała żywiciela, np. tkanki, narządy wewnętrzne.
Kręgowce dostarczają pasożytom nie tylko pokarmu, ale przede wszystkim umożliwiają im rozmnażanie się i ekspansję. Pasożyty mogą nawet zmieniać zachowania swoich żywicieli (np. osobniki są bardziej ruchliwe, skłonne do kopulacji), co ułatwia im rozprzestrzenianie się.
Cykle życiowe helmintofauny mogą być:
- prosty – cały rozwój odbywa się w ciele żywiciele, tylko jajo i larwa przebywają przez krótki okres czasu w środowisku zewnętrznym
- złożony – różne postacie larwalne występują w różnych żywicielach pośrednich i mogą przebywać też w środowisku zewnętrznym.
Wszystkie kręgowce, w tym płazy, spełniają ważną funkcję w cyklach rozwojowych pasożytów. Fauna pasożytnicza polskich płazów obejmuje różne gatunki przywr digenetycznych (Digenea, Trematoda) i nicieni (Nematoda) oraz jeden gatunek przywry monogenicznej (Monogenea), dwa gatunki tasiemców (Cestoda) i dwa gatunki kolcogłowów (Acanthocephala).
Dużą grupę pasożytów płazów stanowią przywry digenetyczne (Trematoda). Zasiedlają one większość narządów wewnętrznych, np. pęcherz moczowy, jelito cienkie i grube, wątrobę, płuca, gonady.
Diplodiscus subclavatus znajdowany jest w końcowym odcinku jelita cienkiego i w jelicie grubym. W wodzie z jaj wydostają się ruchliwe larwy miracidia, które po wniknięciu do ślimaka zatoczka pospolitego (Planorbis planorbis) przekształcają się w cerkarie. Następnie cerkarie opuszczają żywiciela pośredniego i przyczepiają się do kamieni, skorupek ślimaków na dnie stawu. Następnym etapem jest wwiercenie się w skórę żaby i powstanie cyst. Płaz podczas linienia zjada zrzuconą skórę i przenosi cysty do jelita, gdzie uwalniane są pasożyty. Prawdopodobnie do zarażenia dochodzi zimą podczas hibernacji płazów na dnie zbiorników.
Obecność pasożytów i żywicieli w tym samym siedlisku osiąga wysoki stopień dopasowania. Cerkarie Opisthioglyphe rastellus (Fot.1.) po opuszczeniu ślimaka (Galba palustris) opadają na dno zbiornika wodnego. Nastepnie kijanki żaby trawnej (Rana temporaria) żerując przy dnie wywołują wibracje wody. Ruch ten jest sygnałem dla cerkarii do płynięcia w kierunku kijanek. Pasożyty wnikają przez otwór gębowy i skrzelowy do jamy gębowej i tworzą cysty w nabłonku. Po kilku dniach odpadają i zostają połknięte przez płaza.
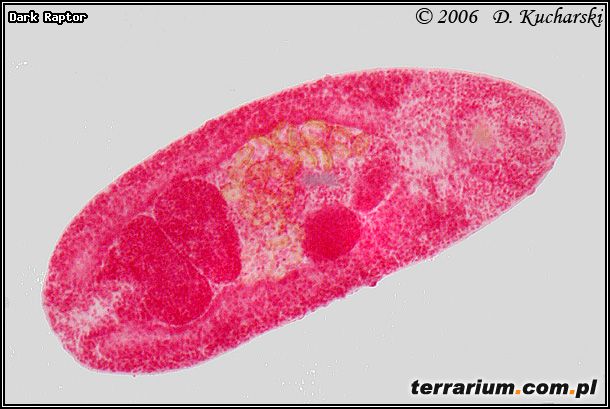
Fot.1. Opisthioglyphe rastellus.
Z kolei u innej przywry Opisthioglyphe ranae cerkarie po wydostaniu się z błotniarki stawowej (Lymnaea stagnalis) płyną pod powierzchnię lustra wody. Gdy żaba porusza podgardlem wywołuje drgania wody. Sygnał ten uaktywnia cerkarie, które docierają do jamy gębowej i przełyku płaza. W nabłonku jamy gębowej otaczają się cystą i są połykane przez żabę. W jelicie odbywa się dalszy rozwój pasożytów.
Pasożytem o złożonym cyklu rozwojowym jest Haematoloechus variegatus (Fot.2.) zasiedlający płuca. Żywicielami pośrednimi tego gatunku są ślimaki zatoczki ostrokrawędziste (Anisus vortex) i larwy komarów (Anopheles maculipennis).
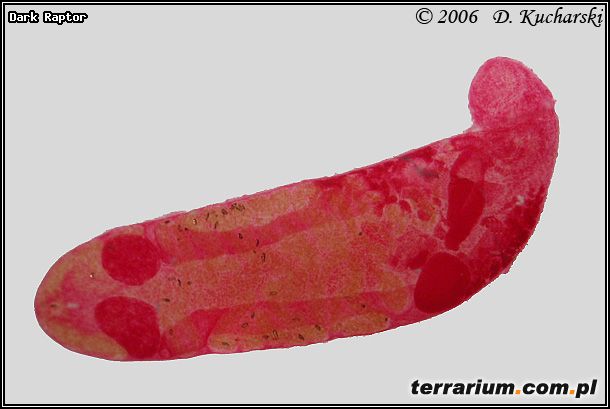
Fot.2. Haematoloechus variegatus.
Innymi przywrami (Trematoda) spotykanymi często u płazów są: Gorgodera cygnoides (pęcherz moczowy), Codonocephalus urniger (gardziel, płuca, wątroba, gonady, uk. moczowy) (Fot.3.), Cathaemasia hians (jelito grube), Pleurogenes claviger (jelito cienkie) (Fot.4.), Pleurogenoides medians (jelito cienkie) (Fot.5.), Prosotocus confusus (jelito cienkie) (Fot.6.), Strigea sp. (jama otrzewnowa), Halipegus ovocaudatus (gardziel, żołądek) (Fot.7.), Haematoloechus coloradensis (płuca Rana pipiens, Ambystoma tigrinum), cysty – formy przetrwalnikowe (gonady, jama otrzewnowa, śledziona) (Fot. 8 ).
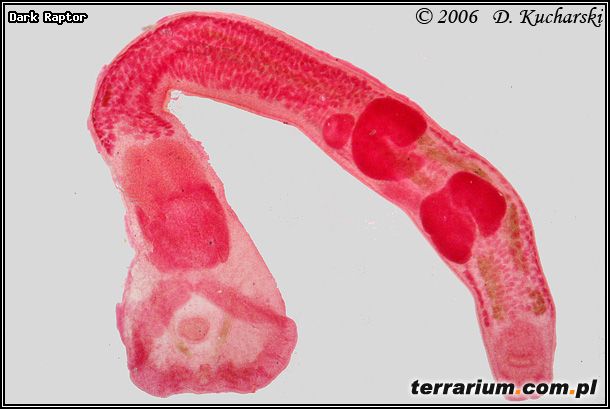
Fot.3. Codonocephalus urniger.
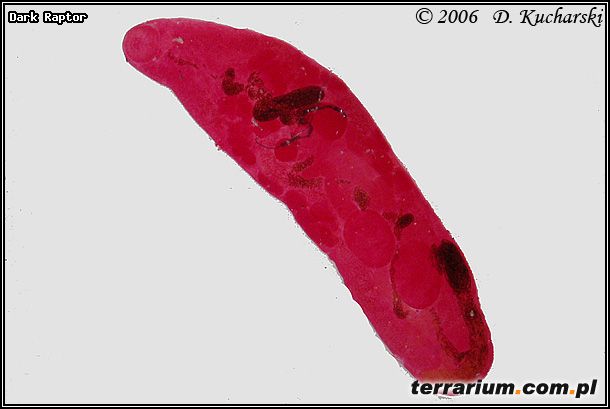
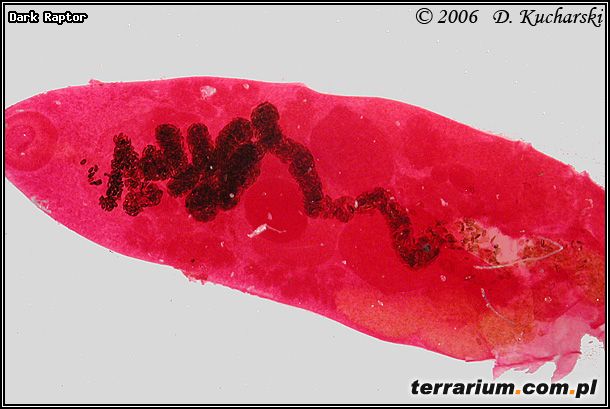
Fot.4. Pleurogenes claviger.
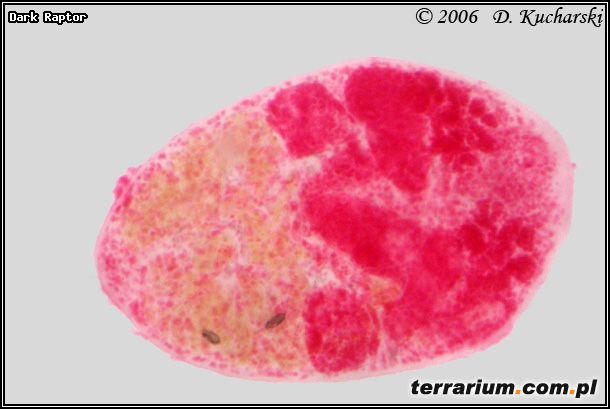
Fot.5. Pleurogenoides medians.
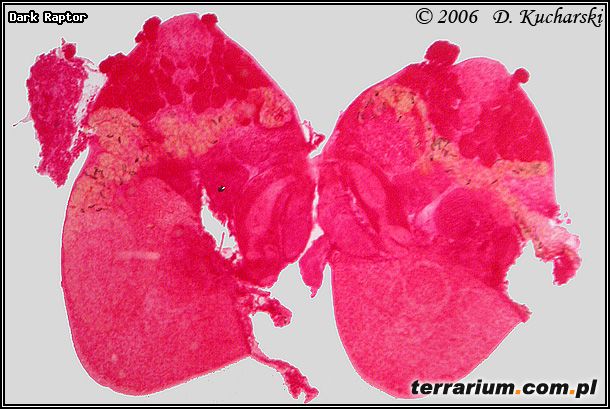
Fot.6. Prosotocus confusus.
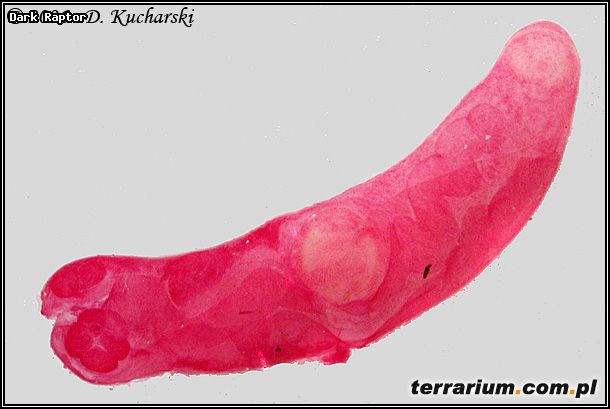
Fot.7. Halipegus ovocaudatus.
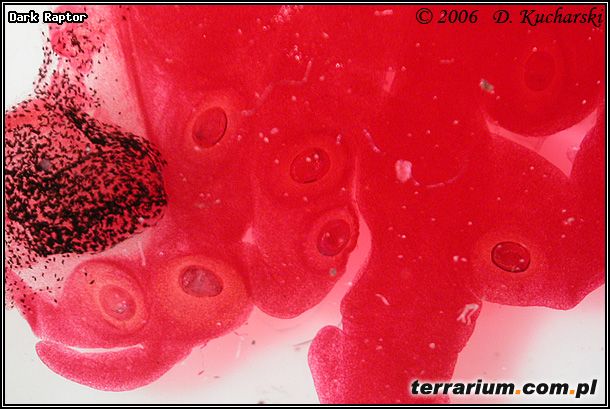
Fot.8. Cysty przywr.
Rozwój prosty, bez przeobrażeń występuje u Polystoma integerrimum (Monogenea). Dorosłe przywry pasożytują w pęcherzu moczowym.
Do nicieni (Nematoda) często spotykanych w jelicie cienkim płazów należą gatunki z rzędu Strongylida: Oswaldocruzia filiformis i z rzędu Ascaridiidae: Cosmocerca ornata (jelito cienkie i grube) (Fot.9.).

Fot.9. Nematoda.
Pasożyty mogą wpływać na zachowania i wygląd swoich żywicieli pośrednich, co ułatwia zjedzenie zarażonego żywiciela. Larwy kolcogłowa Acanthocephalus ranae (Fot.10.) powodują, że ośliczka wodna (Asellus aquaticus) jest bardziej ruchliwa i przemieszcza się w kierunku tafli wody, stając się tym samym podatniejsza na atak płaza. Niezarażone ośliczki przebywają na dnie zbiorników wodnych. Żywicielami ostatecznymi Acanthocephalus ranae mogą być ropuchy, żaby i kumaki.
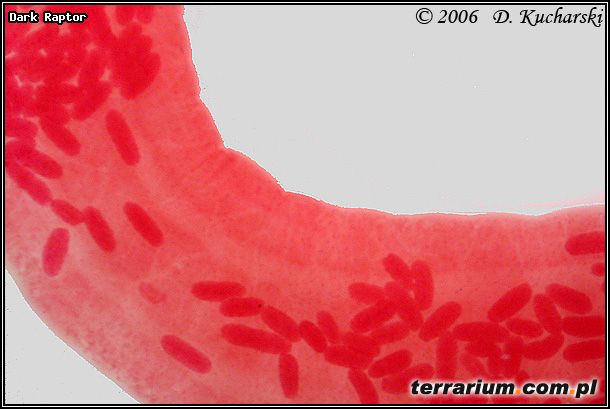

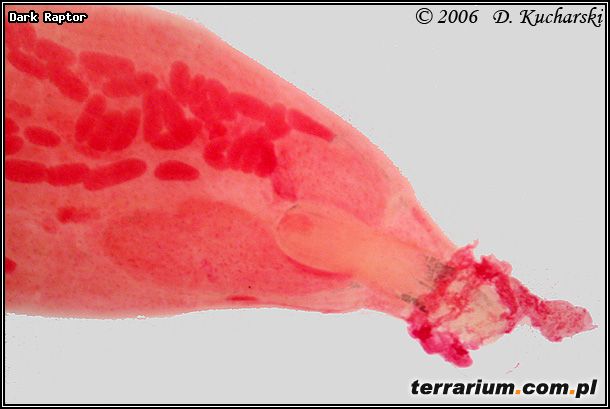
Fot.10. Acanthocephalus ranae.
Drugim gatunkiem kolcogłowa występującego u płazów jest Acanthocephalus falcatus (gatunek górski).
Przykładem pokazującym stopień zapasożycenia dziko żyjącej populacji krajowych płazów może być badanie osobników Rana esculenta complex (n=5) przeprowadzone w czerwcu 2004 r. nad J. Łuknajno. Tabela pokazuje skumulowaną liczbę pasożytów w populacji żab (Tab.1.). Gatunkiem osiągającym największą liczebność jest Codonocephalus urniger (333 osobniki). W ciele jednego z osobników znaleziono 276 tych pasożytów (tzw. efekt „over dispertion” – część osobników jest bardziej podatna na zapasożycenie, np. może to być uwarunkowane genetycznie).
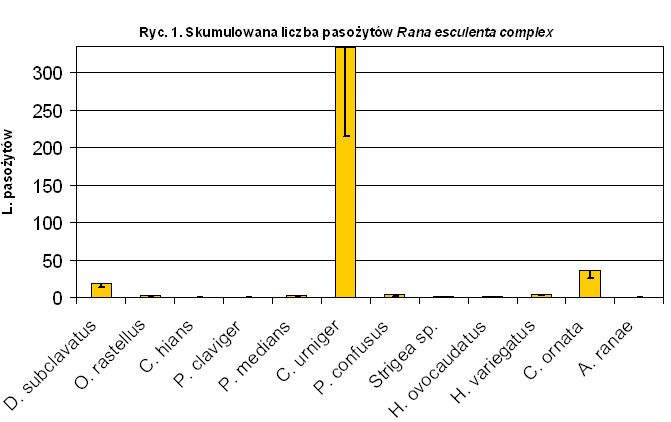
Tab.1. (Według D. Kucharski i B. Makowski).
Obecnie liczebność populacji różnych płazów drastycznie spada. Związane jest to głównie z przekształceniami środowiska naturalnego, co prowadzi do osłabienia osobników, czyniąc je bardziej podatnymi na inwazje helmintofauny.
Układ pasożyt – żywiciel może istnieć gdy spełnione są określone warunki:
1. pasożyt i żywiciel muszą się spotkać w środowisku, czyli muszą przebywać razem w tym samym miejscu i w tym samym czasie (czynnik ekologiczny).
2. tryb życia obu organizmów musi być podobny, co umożliwia bezpośredni kontakt (czynnik etologiczny).
3. żywiciel musi spełniać wymagania pokarmowe pasożyta (czynnik metaboliczny).
4. partnerzy muszą się wzajemnie akceptować (żaden nie powinien uśmiercać drugiego) (czynnik immunologiczny).
Pokonanie tych barier umożliwia skolonizowanie żywiciela. Czas trwania związku pasożyt – żywiciel można wpisać w „ewolucyjny wyścig zbrojeń”. Żywiciel musi wykształcić mechanizmy ograniczające kolonizację dużej ilości helmintofauny, a pasożyty muszą być odporne na te mechanizmy. Przeżywają tylko zwierzęta najlepiej przystosowane, najsilniejsze. Wyścig zbrojeń trwa nieprzerwanie (Hipoteza Czerwonej Królowej – określenie zapożyczone z książki L. Carrolla „Przygody Alicji po drugiej stronie lustra”. Czerwona Królowa powiedziała do Alicji: „musisz biec tak szybko jak tylko możesz, aby pozostać w tym samym miejscu”). Uważa się również, że powstanie płci związane jest ze zjawiskiem pasożytnictwa. Koszty rozmnażania płciowego są wysokie w porównaniu z kosztami rozmnażania bezpłciowego (bez udziału gamet, np. przez podział komórki). Nowe pokolenie osobników powstałe w wyniku namnażania (rozmnażanie bezpłciowe) nie jest zróżnicowane genetycznie (nie pojawiają się np. nowe przystosowania). Z kolei płeć warunkuje rozmnażanie płciowe (gamety męskie i żeńskie) i związaną z tym różnorodność genetyczną. Dzięki niej żywiciele mogą nabywać nowe cechy, ułatwiające im unikanie pasożytów (np. większą odporność).
Opracowanie i źródła informacji
Kornelia Orczyk na podstawie literatury:
Grabda – Kazubska B. 1972 Pasożyty płazów i gadów, PWN
Lonc E., Złotorzycka J. 1994 Zajęcia praktyczne z parazytologii dla studentów biologii, Wydawnictwo Uniwersytetu Wrocławskiego, Wrocław
Lonc E., Złotorzycka J. 1995 Ćwiczenia z parazytologii dla studentów biologii, Wydawnictwo Uniwersytetu Wrocławskiego, Wrocław
Mackenzie A., Ball A.S., Virdee S. R. 2000 Dynamika układu pasożyt – żywiciel, Krótkie wykłady – Ekologia, Warszawa Niewiadomska K. 2005 Jak pasożyty opanowują swoich żywicieli? Kosmos T.54 (2005) Nr 1 (266)
Niewiadomska i wsp. 1981 Zarys parazytologii ogólnej, PWN
Pojmańska T. 2005 Pasożytnictwo, pasożyty i żywiciele. Kosmos T.54 (2005) Nr 1 (266)
Stefański W. 1963 Parazytologia weterynaryjna, tom I, PWRiL, Warszawa
Liczba wyświetleń: 3942


















