




















 [18]
[18]


Chelus fimbriata – źółw matamata
Chociaż matamata (Chelus fimbriata Schneider 1783) jest bardzo znanym żółwiem, nadal jest bardzo mało zbadany. Jego niepowtarzalny, dziwaczny wygląd sprawia, że jest łatwo rozpoznawalny nie tylko dla specjalistów i hobbystów. Gatunek ten jednak stosunkowo rzadko trzymany jest w domach. Ogromne rozmiary akwaterrarium niezbędnego do chowu tego żółwia często przekraczają możliwości normalnych mieszkań. Ponadto, o ile utrzymanie tego gatunku jest relatywnie proste, sukces reprodukcyjny jest rzadko spotykany.
Żółw matamata (Chelus fimbriata Schneider 1783) opisany został ponad 250 lat temu. Przed stworzeniem dwuczłonowej nazwy takson ten funkcjonował jako Testudo terrestris major putamine echinato et striato (Barrere 1741) Prawdopodobny rejon jego występowania określano jako lokalizację „Cayenne” w Gujanie Francuskiej. Nazwę tę następnie skrócono do Testudo terrestris, którą w 1963 roku odrzuciło ICZN (International Commission on Zoological Nomenclature), jednocześnie ustalając nazwę Testudo fumbriata, oraz alternatywne: Testudo matamata i Testudo bispinosa. W 1806 roku gatunek przeniesiono do nowego rodzaju Chelus (łac.) czy Chelys (niem.). W 1890 w wyniku pomyłki powstała kolejna nazwa Chelys boulengeri, która została oparta na podstawie analizy przypadkowej czaszki żółwia z zachowanym zrogowaciałym dziobem (rhamhotheca), podczas gdy wcześniejsze opisy nie uwzględniały tego elementu (w rzeczywistości wszystkie matamaty go posiadają, jednak cecha ta nie jest tak widoczna jak w przypadku innych gatunków żółwi). Nazwy Matamata jako takiej, użyto po raz pierwszy w 1820 roku.
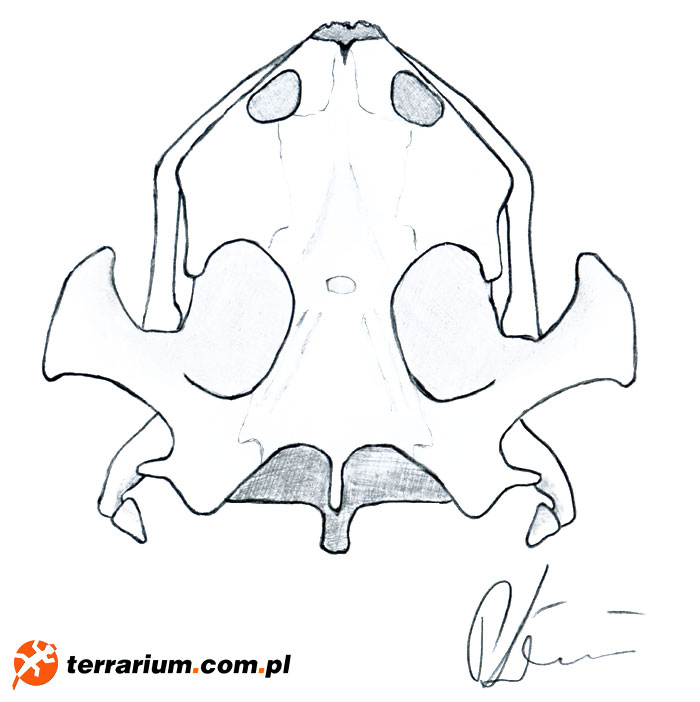
Nazewnictwo
Nazwa polska: Żółw Matamata, Matamata
Nazwa angielska: Matamata turtle
Nazwa niemiecka: Senschildkröte, Mata-Mata
Nazwa czeska: Matamata třásnitá
Nazwa słowacka: Matamata strapcavá
Synonimy
Testudo terestris, Testudo fimbriata, Chelus fimbriata, Chelys fimbriata, Matamata fimbriata, Testudo fimbria, Testudo matamata, , Testudo Rapara, Testudo Raparara, Testudo Raxarara, Emydes matamata, Chelus matamata, Chelys matamata, Testudo bispinosa, Chelys bispinosa, Matamata bispinosa, Chelys boulengerii.
Występowanie
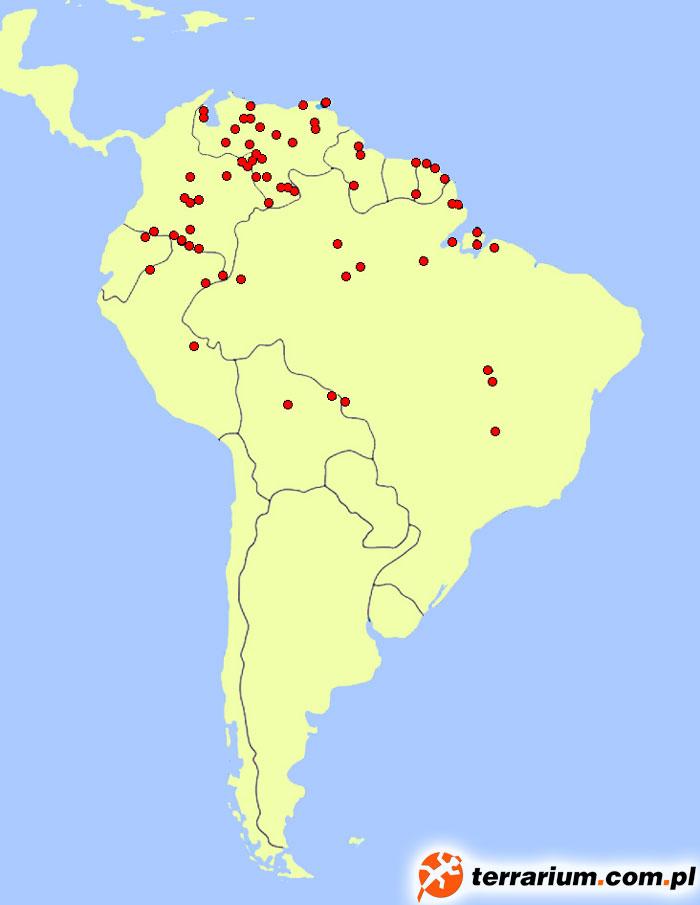
Matamata wydaje się być całkiem licznym gatunkiem, jednak z racji ich skrytego trybu życia, ich ostateczna liczba może być mocno niedoszacowana. Na intensywniej monitorowanych odcinkach rzek Casiquiare i Rio Negro w rejonie wenezuelskiego Llanos są one dobrze znane przez okoliczne plemiona – dla wielu Indian żółwie i ich jaja stanowią urozmaicenie diety.
Żółw matamata występuje na terenach Kolumbii, Wenezueli, Gujany, Gujany Francuskiej, Brazylii, Ekwadoru, Peru, Boliwii, a także wyspę Trynidad. Ściślej ujmując zasiedla on większą część (być może całość) nizinnego sytemu rzecznego Orinoko i Amazonki, a także środkowe i górne Essequibo (Gujana). Występuje na terenach zalewowych Amazonii oraz w ujściu Amazonki, na zachód w obszarze wyspy Marajó przez stan Amapá (Brazylia) i rzekę Oyapoque (wschodnia Gujana Francuska) aż po Cayenne. Żółwie znajdowane na zachodzie Oyapoque mogą być okazami przyniesionymi przez prąd i nie rozmnażać się, podobnie jak te odnajdowane u wybrzeży Trynidadu. W przypadku okazów z Cayenne szacuje się, iż wiele z nich mogło zostać tam przyniesione przez człowieka z Brazylii czy Gujany Francuskiej. W Gujanie Francuskiej matamaty występują na wschodzie w rzekach Oyapock i Approuague, na bagnach Kaw, rzadziej w rzekach Kourou, Sinnamary, Mana. Prawdopodobnie gatunek ten nie jest obecny w Surinamie (jest tylko jeden stary i wątpliwej wiarygodności raport stwierdzający tam jego występowanie z 1878 roku). Szacuje się także, że żółw występuje w jeziorze Maracaibo w wyniku ingerencji człowieka.
Wygląd
Rodzaj Chelus jest przedstawicielem Pleurodira (żółwi bokoszyjnych). Charakteryzuje się silnym bocznym wygięciem kręgosłupa na odcinku szyjnym, obecnością płytki intergularnej i brakiem kości kwadratowo-jarzmowej.
Matamata jest wysoce wyspecjalizowanym taksonem, a jego związek z rodziną (Chelidae) do której należy nie jest oczywisty. Różni autorzy przedstawiają dowody świadczące o możliwie najbliższym pokrewieństwie z rodzajem Hydromedusa lub Phrynops oraz nieco dalszym z rodzajem Chelodina.
Opisano dwie mioceńskie skamieniałości rodzaju Chelus: Chelus colombianus oraz Chelus lewisi odnalezione w Wenezueli. Oba taksony były nieco większe od współczesnego – przeciętnie mierzyły 50, w wyjątkowych przypadkach nawet 60 cm. Niektórzy autorzy nie uznają różnic między tymi gatunkami kierując się wysokim zróżnicowaniem współcześnie żyjącego przedstawiciela rodzaju Chelus.
Pancerz żółwia jest szeroki i niski (zwykle owalny bądź wydłużony), z niezwykle szorstką, urzeźbioną powierzchnią (za wyjątkiem najstarszych osobników). Owalny karapaks zdobią trzy wydatne, guzowate i stożkowate kile, z których każdy osiąga szczyt na linii płytek wertebralnych (przy/okołokręgosłupowych) bądź kostalnych. Na czwartej płytce wertebralnej znajduje się najwyższy punkt karapaksu. Tarczka karkowa jest szeroka, urzeźbiona o kształcie klepsydry lub równoległa. Pierwsza płytka wertebralna jest znacznie szersza od pozostałych. a ich szerokość zmniejsza się aż do najmniejszej nr 5. Brzeg karapaksu jest falisty bądź ząbkowany, szczególnie w części tylnej. Pancerz żółwia jest raczej spłaszczony niż zbieżny, otwierając się szeroko w przedniej części („wlot muszli”). Most jest krótki (stanowi zaledwie ok. 25% długości plastronu – jest znacznie krótszy niż u skamieniałych przodków Chelus). Na plastronie krótki most koresponduje ze zwykle krótkim szwem interabdominalnym. Płytka intergularna jest bardzo zróżnicowana pod względem kształtu i wielkości i może choć nie musi występować między płytkami gularnymi. Doniesiono także iż okazjonalnie może występować pośrodkowo mała płytka tuż za płytką intergularną. Najdłuższym szwem plastronu jest szew występujący między tarczkami femoralnymi. Tylny płat plastronu jest wąski, zwężający się ku tyłowi o niemal prostych bokach. Wcięcie analne jest dobrze rozwinięte, kanciaste u osobników młodocianych i zaokrąglone u dorosłych. Kolor karapaksu jest zmienny, począwszy od jasnobrązowego u młodych osobników, ciemnobrązowego u dorosłych, aż po niemal czarny u najstarszych żółwi. Płaska i trójkątna głowa jest niezwykle szeroka i spłaszczona z nozdrzami umieszczonymi na szczycie wyraźnej, wąskiej podwójnie-cylindrycznej mięsistej rurki (trąbkowatego nosa). Głowa, wraz z szyją pokryta jest szeregiem fałd skórnych o rozmaitym kształcie i fakturze, co wraz z mnóstwem guzków i wyrośli daje to obraz prehistorycznego dziwadła. Oczy są małe i umieszczone w przedniej części. Za oczami znajduje się szeroki płat skóry. Otwór gębowy jest bardzo szeroki. Potężna wystająca paszcza pozwala przypuszczać jak duże kęsy zwierze może połknąć. Występują dwie krótkie brodawki na dolnej szczęce dokładnie pod orbitami oczu oraz poprzeczne serie złożone z 4 płatów skóry na szyi biegnące na wysokości błon bębenkowych. Szyja jest długa oraz bardzo gruba i muskularna z szeregami płaskich, rozgałęzionych płatów skórnych wzdłuż jej bocznej krawędzi. Spodnia część szyi może być od brązowej po czerwoną, z ciemnobrązowymi, bądź zwłaszcza u młodych osobników czarnymi pasami. Na wierzchniej stronie szyi może pojawić się strzałkowaty wzór koloru czarnego. Głowa wraz z szyją mają w przybliżeniu tą sama długość co karapaks. Kończyny są raczej małe i słabe o stosunkowo małych palcach spiętych niewielką błoną. Jest po 5 pazurów na każdej przedniej łapie oraz po 4 na tylnych. Kości karapaksu są umiarkowanie grube zwłaszcza wzdłuż grzbietu.
Odmiany
Duży zasięg występowania żółwi matamata pozwala przypuszczać że występują różne formy geograficzne. Początkowo nie wyodrębniono żadnych podgatunków, jednak w 1966 stwierdzono różnice w obrębie odrębnych populacji matamat, tym samym charakteryzując formy terytorialne. Różnice te dotyczyły żółwi pochodzących z Kolumbii, Peru oraz Brazylii, jednak prawdopodobnie porównywano jedynie osobniki zasiedlające Amazonkę. Podstawowe różnice stwierdzono w obrębie koloru karapaksu i plastronu, głowy, tarczki karkowe, faktury karapaksu, kształtu czaszki, płatów skórnych nad uchem, płytki intergularnej.
Badania większej grupy żółwi pochodzących z Orinoko oraz Amazonki wykazały liczne różnice i na ich podstawie dokonano typologicznego rozróżnienia na dwie odmiany terytorialne. Te pochodzące z Orinoko miały owalne lub jajowate skorupy oraz mało lub wcale czarnego barwnika po spodniej stronie szyi (zwykle były jasnoróżowe). Te pochodzące z Amazonki miały karapaks wydłużony lub nawet zwężony, prostokątny z dwoma szerokimi ciemnymi paskami biegnącymi po spodniej stronie szyi, które rozjaśniają się pod żuchwą. Charakterystyczna kolorystyka szyi i plastronu opisana powyżej, bardzo ułatwia rozróżnienie zwierząt młodych. U dorosłych osobników najłatwiej rozpoznać odmianę po kształcie karapaksu. Również kolorystyka jest różna, typ amazoński ma plastron brązowy do ciemnobrązowego z jasnoczerwonymi plamkami, natomiast Orinoko bardziej żółtawy, jasnobrązowy lub w kolorze ochry z kilkoma ciemnymi plamkami. Niektórzy autorzy stwierdzają ogólnie, że plastron dorosłych żółwi z Orinoko nie posiada pigmentu w odróżnieniu do tych zasiedlających wody Amazonki.
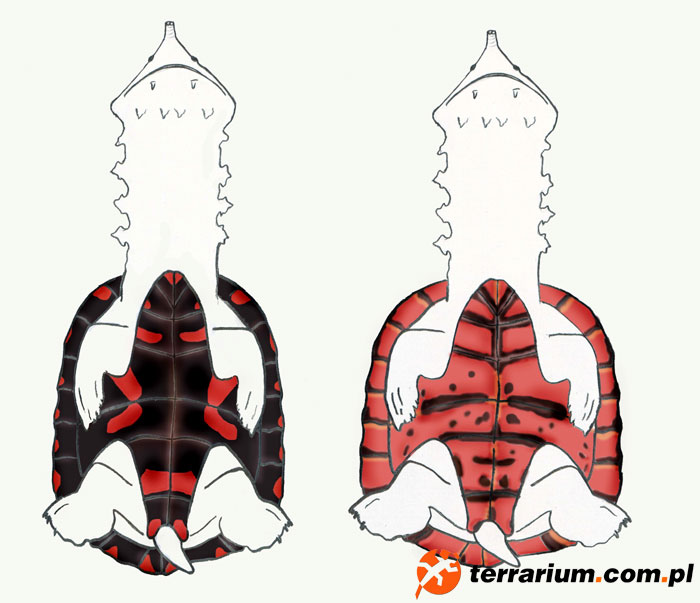
Te dane sugerują występowanie dwóch podgatunków Chelus fimbriata: jednego w Orinoko i Essequibo (Kolumbia, Wenezuela i Gujana) oraz drugiego w Amazonce (Brazylia, Peru, Kolumbia, Ekwador, Boliwia oraz region Oyapoque w Gujanie Francuskiej). Styk obu populacji może mieć miejsce w górnym odcinku Rio Negro (Wenezuela, Kolumbia i Brazylia).
Jednakowoż podobne badania prowadzone na populacji Wenezuelskiej ujawniły dużą zmienność osobniczą wewnątrz małej grupy 12 żółwi. Wyjątki odnoszące się do ubarwienia tych form terytorialnych odnoszą się do egzemplarzy zasiedlających rejony mieszania się obu populacji (Rio Negro). Jeżeli chodzi o posiadanie bądź brak płytki intergularnej udowodniono, że w obu populacjach procent jej występowania jest niemal identyczny.
Genetyczne badania powinny zasadniczo pomóc w określeniu czy w przypadku tego gatunku mamy do czynienia z jednym mocno zróżnicowanym taksonem, czy też oddzielnymi podgatunkami.
Matamata w jeziorze Maracaibo
Jest to największe jezioro w Ameryce Południowej . Różni autorzy wspominali o możliwości występowania tam żółwi matamata, jednak nie potwierdzono tej wiadomości. Warunki jakie oferuje zbiornik byłyby bardzo dogodne dla żółwi matamata. Szacuje się, że gdyby te gady zostały tam odnalezione należałoby poddać je badaniom z uwagi na możliwość oznaczenia nowego podgatunku bądź nawet gatunku lub taksonu znanego jedynie ze szczątków kopalnych jak Chelus lewisi (z Proto-Orinoko w Formacji Urumaco w północno-zachodniej Wenezueli).
Uwaga
Jakkolwiek żółwie importowane z Orinoko cieszą się dużym powodzeniem to należy podkreślić, że z wiekiem ich kolorystyka traci na atrakcyjności. Plastron staje się jednolity i jasny (cielisty do kremowego). Mimo iż jest to odmiana najczęściej jaskrawo ubarwiona na spodniej stronie ciała to osobniki z Amazonki są bardziej kontrastowe zarówno jako młode jak i dorosłe egzemplarze. Na uwagę zasługuje także fakt, że w kolekcjach i hodowlach amatorskich częściej spotykaną formą tego żółwia są właśnie zwierzęta w typie Orinoko.
Wielkość i tempo przyrastania
Matamata jest prawdopodobnie największym przedstawicielem swojej rodziny (na pewno w Ameryce Południowej), zważywszy na przeciętne rozmiary dorosłych egzemplarzy, choć godnymi rywalami są australijska Chelodina expansa i Elusor macrurus. Podczas gdy dorosłe samce rzadko osiągają 40 cm CL (długości karapaksu), dorosłe samice zwykle przekraczają tę długość. Początkowe badania rozmiarów tych żółwi (1976 rok) wykazały zakres od 31.1 cm do 40.4 cm CL przy grupie 19 nieseksowanych dorosłych egzemplarzy (prawdopodobnie większość z nich była samcami). Wielu badaczy nie zdawało sobie sprawy jak duże mogą być dorosłe samice. W 1977 roku określono dwie z nich długości 36,5 i 39,8 cm CL jako wyjątkowo duże i bardzo stare, oraz jedną samicę długości 44,9 cm jako niezwykle dużą i bardzo rzadko spotykaną. Podobnie orzeczono w przypadku samicy odłowionej w tym samym roku w Peru – mierzyła ona 43,7 cm CL. Dziś wiadomo, że wymiary te są średnie dla dorosłych samic. Niektórzy autorzy przesadnie szacowali maksymalne wymiary tych żółwi („długość około jednego jarda”) dając im maksymalnie 125 cm CL oraz 225 cm długości całkowitej! Największy okaz zmierzony w 1984 roku z Puerto Ordaz w Wenezueli mierzył 46,1 cm CL, natomiast największy z grupy 105 egzemplarzy poddanych badaniu w 1995 roku miał 46,0 cm CL. Wymiary te zostały pobite przez największego z 15 dorosłych żółwi z Gujany, który miał maksymalną długość CL 46,4 cm i masę 11,45 kg; najcięższym z nich był jednak 44,7 centymetrowa samica o masie 11,70 kg. W 1999 roku doniesiono o matamacie z Puerto Ayacucho (Wenezuela) o CL 48,2 cm (według innych źródeł 49 cm, przy czym był to jedynie sam pancerz). Badania opublikowane w 2006 roku wykazały samicę z Río Apure o CL 52,6 cm (przy linii środkowej długości 50,2 cm) i masie 17,7 kg – okaz ten został z powrotem wypuszczony. Naukowcy dyskutują na temat pomiarów długości pancerza matamaty, stąd możemy wnioskować, że CL nie określa precyzyjnie długości skorupy żółwia. Dokonanie pomiaru po krzywiźnie jest oczywiście możliwe choć pancerz matamaty jest na tyle płaski, że ten sposób pomiaru mógłby dodać najwyżej parę centymetrów. Żółw posiada niezwykle dużą czaszkę, szerokości do 15,0 cm (okaz z San Carlos de Río Negro, Wenezuela – według proporcji żółw ten winien mieć 52,8 cm długości). W badaniach prowadzonych na grupie 10 dorosłych samic z Orinoko przy średniej CL rzędu 43,1 cm wykazały średnią szerokość czaszki 12,25 cm (maksimum 13,0 cm dla CL 45,0 cm). Porównując te wymiary z 9 dorosłymi samicami z południowej Gujany o średniej długości CL 44,25 cm i szerokości czaszki 12,01 cm można szacować iż długość CL dla okazu o szerokości czaszki 15,0 cm mogła wynosić 55,27 cm – wielkości przystającej kopalnym przodkom jak C. colombianus czy największym C. lewisi.
Na Uniwersytecie w Wenezueli (Universidad Simon Bólivar, Sartenejas) prowadzono kompleksowe badania dotyczące zwłaszcza morfologii (cech zewnętrznych) obu przedstawicieli rodzaju Chelus (wymarłego i żyjącego współcześnie).
Dwanaście dorosłych samców z Gujany miało średnio 39,2 cm CL (37,4 – 41,9 cm), podczas gdy 10 samic mierzyło średnio 44,4 cm CL. Przeciętna relatywna szerokość karapaksu – CW (ang. carapace width) w przypadku obu wymienionych grup była praktycznie identyczna; CL/CW 1,307 dla samców oraz 1,298 dla samic.
W badaniach 14 samców z Rancho La Trinidad, Estado Cojedes w Wenezueli średnia długość CL wyniosła 35,8 cm (31,1 – 39,6 cm), 9 samic natomiast przeciętnie 43,2 cm (40,8 – 45,0 cm).
W obu przypadkach wymienionych wyżej grup samce były mniejsze od samic i bardziej liczne, co potwierdza pogląd, że w gatunku tym płeć osiągająca dojrzałość wcześniej nie tylko pozostaje mniejsza w stosunku do płci przeciwnej, ale także przewyższa ją liczebnie.
Dwa żółwie matamata utrzymywane w warunkach sztucznych urosły od około 8 cm CL (ang. carapax lenght – długość karapaksu) do 25 – 30 cm w przeciągu 9 lat. Okazuje się także, że pierścienie na tarczkach są stosunkowo odmienne w zależności od siedliska, zwłaszcza w przypadku zwierząt zasiedlających wody w obrębie sawann bądź llanos, gdzie dostępność pokarmu jest znaczna w porze suchej, natomiast w porze wilgotnej zwierzęta, którymi odżywia się matamata są znacznie bardziej rozproszone. Stąd sukcesywne polowanie może być na tyle rzadkie by zahamować wzrost żółwia. Badania tarczek okazów pochodzących z Orinoko dowiodły, że egzemplarz mierzący 28.1 cm CL był w wieku 5 lat, podczas gdy inny długości 18,6 cm liczył 5,5 roku. Wiadomo zatem, że tempo wzrostu żółwi jest bardzo zróżnicowane. Badania trzech dorosłych samców z południowej Gujany sugerowały, iż osobnik długości 37,4 cm miał 14 lat, przy znacząco wolniejszym wzroście w przeciągu ostatnich dwóch lat; okaz 37,9 cm był w wieku 16 lat z ostatnimi 5 pierścieniami przyrostu bardzo wąskimi (powolny wzrost); 37,5 cm żółw był w wieku 11 lat bez znaków spowolnionego wzrostu. W trakcie tych samych badań oszacowano wiek innych żółwi:
-
- 10 lat (38,8 cm);
-
- 11 lat (39,1 cm);
-
- 12 lat (38,0 cm);
-
- 15 lat (40,0 cm);
-
- 18 lat (40,8 cm).
Cztery samice z policzalnymi pierścieniami przyrostów oznaczono na:
-
- 13 lat (43,8 cm);
-
- 15 lat (44,1 cm);
-
- 20 lat (43,5 cm);
-
- 20 lat (46,4 cm).
Biotop
Matamata określany jest jako gatunek wybitnie wodny, nizinny i tropikalny. Znajdowany jest zarówno na terenach gęsto zalesionych jak i w rzekach powiązanych z sawannami jak Rupununi w Gujanie czy llanos (trawiasta formacja roślinna, charakterystyczna dla sawanny w Ameryce Południowej) w Wenezueli. Gatunek zasiedla głównie systemy rzeczne Amazonki i Orinoko, w systemie Essequibo (Gujana) pojawia się głównie powyżej bystrza i zaczyna być ograniczony w środkowej części oraz jest nieobecny blisko wybrzeża. Matamata występuje w bardzo zróżnicowanych habitatach wodnych, ale preferuje starorzecza, ciche stawy oraz relatywnie małe zatoki o łagodnym i powolnym przepływie wody. Jednakowoż żółwie są regularnie (przypuszczalnie biernie) przenoszone z ujścia Orinoko i czasem odnajdowane jako żywe wyrzucone na południowym wybrzeżu Trynidadu. Obecność organizmów morskich na muszlach żółwi świadczy o fakcie że były w stanie przetrwać w warunkach morskich przez okres kilku tygodni. Prawdopodobnie właśnie na skutek takiego transportu żółwie były w stanie skolonizować rzekę Essequibo w Gujanie. Żółwie zbierane w Kolumbii za dnia znajdowano w błocie przyległym do stromego brzegu llanos, w nocy napotykano je kroczące po dnie. Zwierzęta były najbardziej aktywne w bezksiężycowe noce. Młode zasiedlają miejsca w których występują płytkie wiry, zapewniające im ochronę, dostępność piaszczystego substratu oraz opadłe liście do których są podobne. Matamaty są raczej słabymi pływakami i unikają prądu rzecznego zajmując refugia (schronienia) pod gałęziami, pniami i kłodami zanurzonymi w wodzie przy brzegu.
Właściwości fizjologiczne i ekologiczne
Matamata jest gadem wybitnie wyspecjalizowanym pod kątem pokarmu. Rozmaite, nieco dziwne modyfikacje jego głowy oraz szyi, są przejawem jego niezwykłego przystosowania względem detekcji i połknięcia ruszającej się ofiary (małej ryby) w warunkach w których widoczność jest niewielka bądź zerowa jak w nocy czy nieprzezroczystej mulistej wodzie. Przystosowania te obejmują klapy skórne oraz liczne frędzle w rejonie głowy i szyi które są intensywnie unerwione i poruszają się pasywnie w odpowiedzi na drobne zakłócenia wody wokół głowy. Niektórzy autorzy twierdzą nawet, że struktury te mają za zadanie wabić ryby. Niewątpliwie jednak spełniają one rolę czułego narządu odbierającego bodźce. Zmierzono potencjał elektryczny tychże wyrostków wystawionych na działanie bodźców, okazało się, że są one na tyle czułe, że odbierają najmniejsze różnice w cyrkulacji wody i ruch niewielkich obiektów w pobliżu głowy żółwia. Zwierzęta te zamieszkują mętne wody, o bardzo słabej widzialności, można więc ze sporą dozą prawdopodobieństwa założyć, że owe wyrostki stanowią niezwykle ważny element budowy umożliwiający detekcje ofiar w najbliższym położeniu. Ponadto rozmiar i szeroka separacja błon bębenkowych świadczą o dalszej wrażliwości żółwia na podwodne wibracje, razem ze zdolnością do wykrywania kierunku takich zakłóceń. Silne spłaszczenie głowy prawdopodobnie ułatwia szybkie, boczne uderzenie zwierzęcia w ofiarę przy jednoczesnym bardzo szerokim otworzeniu i zamknięciu niesamowicie szerokiej paszczy oraz mocnemu naporowi wody (wraz z ofiarą) generowanej przez uderzenie masywnego aparatu gnykowego i przerośniętego (hypertroficznego) mięśnia szyjnego. W płytkiej wodzie wylegujący się matamata, używa do zaczerpnięcia powietrza swojego długiego, elastycznego rurkowatego nosa – żółw wyciąga maksymalnie szyję, a nad powierzchnię wody wynurza się jedynie maleńki koniuszek nosa. Matamata ma małe oczy ale świetny wzrok – tylna część oka jest wysłana jest błona odblaskową która odbija światło. Krokodyle i inne nocne gady również posiadają podobną strukturę.
Przewód pokarmowy żółwi matamata jest niezwykle efektywny i strawiają one znaczącą część spożytego pokarmu, tak więc ilość odchodów którą wydzielają jest relatywnie mała.
Pod względem ekologicznym Chelus fimbriata może pełnić podobną rolę w stosunku do północnoamerykańskiego żółwia sępiego (Macrochelys temminickii). Jest wiele cech łączących oba gatunki, włączając w to dietę złożoną z żywych ryb, strategię łowiecką (leżeć i czekać), duże rozmiary, guzkowaty pancerz oraz zdolność wtapiania się w otoczenie. Oczywiście budowa głowy i paszczy oraz mechanizm łapania i spożywania żywej ofiary różnią się zasadniczo u tych dwóch gatunków.
W typowych warunkach, gdy widoczność bliska jest zeru, zbliżanie się potencjalnej ofiary do głowy żółwia jest przypuszczalnie szczęśliwym zbiegiem okoliczności i zależy raczej od ruchów ofiary niż drapieżnika. W warunkach sztucznych gdzie przejrzystość wody umożliwia wizualne zlokalizowanie ofiary żółw może dosłownie podejść do niej by ją pochwycić. Opisano aktywność matamaty w długim akwarium, który ustawił się tak, że ryba przepływając wzdłuż zbiornika znajdowała się w zakresie uderzeń paszczy żółwia. Obserwowano także „pasterskie” (zaganiające) zachowania matamaty utrzymywanej w wydłużonym, półkolistym basenie, który powoli patrolował całą długość zbiornika z ciałem wygiętym pod takim kątem by blokować większość jego szerokości. Przestrzeń w której znajduje się ryba zmniejsza się stopniowo na jednym końcu zbiornika i jest ona także zmuszona do przepłynięcia obok żółwia co często skutkuje jej sprawnym przez niego schwytaniem.
Utrzymywanie w niewoli
Żółwie te licznie są utrzymywane w warunkach sztucznych, wliczając w to setki żółwi w rękach prywatnych. Już badania z późnych lat 80. XX wieku uwzględniają liczbę 117 sztuk utrzymywanych w niewoli w różnego rodzaju ośrodkach w USA. Z naukowego punktu widzenia chów matamaty nie wymaga żadnych specyficznych warunków, poza zapewnieniem zwierzęciu wody o odpowiedniej głębokości (tak by mogło głową sięgnąć powierzchni krocząc po dnie) oraz żywych ryb do czasu przyzwyczajenia podopiecznego do martwej ofiary bądź jej fragmentów. Matamaty mogą przeżyć temperatury znacznie niższe od tych z jakimi mają do czynienia w naturze, choć temperatura dla nich letalna nie została określona. Doniesienia o pierwszych przypadkach rozmnożenia w niewoli pochodzą z 1943 i 68 roku. Znacznie wcześniej (1835 rok) pojawia się wzmianka o kilku matamatach z Cayenne, w kolekcji muzeum historii naturalnej Panteonu w Paryżu. Jeden z żółwi miał wówczas złożyć jaja i przynajmniej z jednego z nich wykluł się żółw który potem został utrwalony w płynie konserwującym dla celów kolekcji.
Utrzymanie w akwaterrarium
Jeżeli spełnić podstawowe potrzeby utrzymania żółwia matamata, jego chów jest prosty. Podstawowe błędy dotyczą wielkości zbiornika i poziomu wody. Zwierzęta mające dostatek miejsca częściej można obserwować aktywne, a odpowiednio wysoki poziom wody sprawia ze niekiedy nawet pływają pod powierzchnią. Żółwie matamata są stosunkowo mocno podatne na stres i jeżeli dodatkowo trzymane są w akwariach o zbyt małych rozmiarach, może to się odbić na zdrowiu zwierzęcia.
Najodpowiedniejszym podłożem w akwaterrarium jest piasek, młode żółwie często obserwuje się kopiące w piasku, u starszych osobników wydaje się on nie być potrzebny, dlatego wielu hodowców rezygnuje z niego ze względów higienicznych. W przypadku młodych żółwi poziom dna powinien być zróżnicowany, ponieważ zwierzęta nocą wolą przebywać w płytszej wodzie, natomiast na żerowanie i odpoczynek w ciągu dnia chętniej wybierają głębsze partie zbiornika. Zaleca się urządzenie akwaterrarium z wykorzystaniem korzeni i konarów dla zapewnienia żółwiowi schronienia. Część lądowa akwaterrarium wymagana jest jedynie w przypadku dorosłych, przystępujących do rozrodu żółwi. Wielkość części lądowej musi być oczywiście dostosowana do wielkości zwierząt, ale przyjmuje się że dla jednej pary matamata wyspy powinna mieć nie mniej niż 70×50 cm. Podłoże części lądowej musi mieć co najmniej 25 cm grubości, a na czas rozrodu powinna być dodatkowo ogrzewana kablem grzewczym.
Oświetlenie
Chociaż dla żółwia matamata, zwierzęcia prowadzącego w naturze nocny tryb życia, oświetlenie, zwłaszcza światło UV wydaje się mieć marginalne znaczenie, w literaturze spotyka się wzmianki o pozytywnych efektach światła o tej długości fal. Matamata wychodzi z wody jedynie w celu złożenia jaj, nie ma zwyczaju brania kąpieli słonecznych na lądzie. Zmętnienie wody w ich środowisku naturalnym, połączone z gęstą roślinnością, ograniczają niemal zupełnie penetrację promieni słonecznych. Na głębokości 1,5 m nawet w czystej wodzie, widoczność jest bardzo ograniczone przez cień jaki powoduje zadrzewienie brzegów rzeki. Dlatego zwierzęta te najlepiej czują się w zacienionym akwaterrarium i w takim właśnie wykazują największą aktywność wczesnym rankiem i o zmierzchu, natomiast w nocy śpią.
Temperatura
Najczęściej zalecana temperatura optymalna dla matamaty to 28-30°C. Jednak temperatura w jego naturalnym środowisku, mierzona na głębokości od 30 cm do metra w wodzie o słabym nurcie zawiera się w przedziale od 24 do 26°C, natomiast w stojącej, zacienionej wodzie zwykle wynosi poniżej 24 stopni, nawet około 22ºC. Literatura podaje że matamata w temperaturze 17°C nadal się odżywia, jednak temperatura wody zdecydowanie nie powinna spadać poniżej 22ºC. Optymalną temperaturą jest 24-25°C. Zdecydowanie nie poleca się ogrzewania wody w akwaterrarium przez podgrzewanie kablem grzewczym dna zbiornika. Żółwie te spędzając większą cześć życia oparte na dnie, przez co podgrzana szyba może powodować masowe namazanie się niebezpiecznych drobnoustrojów pod powierzchnią plastronu, rezultatem czego są niezwykle trudne do wyleczenia nekrozy.
Uwaga! Niewłaściwa temperatura wody i powietrza w zbiorniku może doprowadzić do poważnych zaburzeń fizjologicznych i behawioralnych u żółwi. Taką sytuację można było obserwować swojego czasu w berlińskim Ogrodzie Zoologicznym. Młode żółwie matamata opuściły zbiornik wodny, wyszły na ląd, część z nich się zakopywała. U podłoża tego fenomenu prawdopodobnie znalazła się przegrzana woda w basenie oraz niska wilgotność powietrza (spowodowana zbyt dużym rozmiarem całego zbiornika w stosunku do skromnego basenu), być może także ogółem nieprawidłowy gradient temperatur.
Woda
Woda w rzekach w których żyją matamaty w większości przypadków jest silnie zakwaszona, miękka i uboga w składniki mineralne. W regionie Amazonki pH wody waha się w granicach 3,7 – 6,1. Na ile stan chemiczny wody ma wpływ na samopoczucie dorosłych matamat trudno jednoznacznie stwierdzić, jednak w przypadku młodych osobników, zauważono, że ograniczają one spożywanie pokarmu gdy woda dłuższy czas nie jest podmieniana. Ilość pobieranych ryb maleje wraz z wzrostem zabrudzenia wody, po czym po podmianie wody wraca znów do pierwotnej ilości. Prawdopodobnie ma to związek ze stężeniem związków azotu powstających podczas rozkładu białka, którego w prawidłowej diecie matamaty jest pod dostatkiem.
Żywienie
Mimo iż wiele uwagi poświęcono mechanizmom polowania matamaty w warunkach sztucznych, praktycznie brak danych na temat składu gatunkowego ryb łapanych przez tego żółwia w naturze, za wyjątkiem najczęściej przezeń chwytanego w Gujanie Francuskiej „characina” (Erythrinus erythrinus) mierzącego najczęściej 20-25 cm długości. Jednak jako, że pobieranie pokarmu u tego gada jest wynikiem szybkiej reakcji na niewidoczny bodziec, prawdopodobne jest, że selekcja ofiar zachodzi jedynie w odniesieniu do przybliżonej oceny ich rozmiarów, które przypuszczalnie mogą być dolnym lub górnym limitem najbliższym organizmowi wywołującemu zachowanie żerowe. Każda ryba zasiedlająca habitat żółwia mieszcząca się w tym zakresie jest potencjalną ofiarą. W warunkach sztucznych żółwie mogą początkowo preferować żywe ofiary, ale szybko dostosowują się do połykania martwych lub nawet ich fragmentów. Nie wiadomo czy w warunkach naturalnych kiedykolwiek spożywają padlinę. Zwyczaje żywieniowe matamaty są specyficzne i często opisywane w literaturze. Ofiara jest zasysana z ogromną prędkością przez raptowne otwarcie ogromnej paszczy aż do szerokiego gardła. Kiedy kęs pożywienia jest już w gardle, albo chociaż trzymany jest mocno w pysku, woda zassana podczas ataku jest wypuszczana przez wąską szczelinę pomiędzy zaciśniętymi szczękami. Matamata jest typowym rybożercą, chociaż raz na jakiś czas przyjmowane są myszy i jednodniowe kurczęta, natomiast w naturze w żołądkach matamat znajdowano kijanki płazów. Każda ryba jest chętnie przez żółwia zjadana, nie należy podawać chorych ryb ozdobnych (zwłaszcza importowanych z habitatu żółwia w Ameryce Południowej), ze względu na możliwość przeniesienia niebezpiecznych patogenów na które żółw będzie wrażliwy. Ryby okazjonalnie mogą być podawane żywe, jednak zwykle zaleca się mrożenie ryb i podawanie ich żółwiowi po rozmrożeniu. Można również od czasu do czasu podawać martwe myszy i kurczęta – jednodniówki. Skarmia się całe ryby, bez oczyszczania. Niektórzy autorzy polecają umieszczanie w podawanych rybach tabletki drożdży piwnych. Raz na dwa – trzy tygodnie w pokarmie podajemy także witaminy. Można również podawać kijanki hodowlanych żab zielonych (Rana esculenta i Rana lessonae), które są chętnie zjadane przez żółwie. Osobnik dorosły powinien otrzymywać, w zależności od pory roku 100 – 300g pożywienia w tygodniu. Poważnym problemem jest otłuszczanie się żółwi trzymanych w terrariach – ważą one średnio 50-70% więcej niż ich pobratymcy o takiej samej średnicy karapaksu żyjący na wolności. Zdarzają się żółwie które żyjąc u nadopiekuńczych właścicieli przekraczają właściwą dla swojej wielkości wagę nawet o 170%. Nadmierne otłuszczenie uważa się za główny problem obniżonej lub zerowej aktywności seksualnej żółwi w niewoli.
Matamaty nie lubią łapać martwych ryb pływających po powierzchni wody, ponieważ wówczas, atakując, nabierają sporo powietrza, stąd obserwowano zwierzęta, w zbiornikach sztucznych, które gdy zobaczyły martwą rybę, pływającą po powierzchni, szybko przenosiły się pod strumień wody wylewanej przez filtr i czekały, aż lejąca się woda wepchnie rybę pod wodę, skąd wygodniej było im ją łapać. Takie zachowanie świadczy o inteligencji i zdolności do szybkiego uczenia się tych zwierząt. [mała grupa badawcza, obserwacje nie potwierdzone eksperymentalnie]
Zachowanie
Matamata to typowo wodny gatunek żółwia, który opuszcza wodę tylko po to żeby złożyć jaja. Żółw prowadzi osiadły, przydenny tryb życia – porusza się częściej krocząc po dnie niż pływając, chociaż notowano dorosłe żółwie w odpowiednio dużych sztucznych zbiornikach, które chętnie pływały. Koresponduje to także faktem częstego ich spotykania w wodach o głębokości po wyżej jednego metra. W naturze obserwuje się żółwie, które unoszą się tuż pod powierzchnią wody w miejscach silniej nasłonecznionych – w ten sposób zwierzęta zażywają kąpieli słonecznych.
W odpowiednio dużym akwaterrarium matamaty stają się aktywne, i zaskakująco zwinne. We wczesnych godzinach porannych można je obserwować pływające i chodzące po dnie. Drugi szczyt aktywności przypada na późne popołudnie i trwa do zapadnięcia zupełnej ciemności (w zależności kiedy będzie miał zgaszone światło).
Kiedy w trakcie tych codziennych aktywności natrafią na siebie dwa samce, wówczas ustawiają się bokiem do siebie, ze zgiętymi na bok szyjami i karapaksem uniesionym od strony rywala próbując zaprezentować większe rozmiary ciała niż w rzeczywistości. Jeżeli dochodzi do drobnych utarczek, żółwie zawsze podgryzają sobie wzajemnie jedynie tylne części ciała (ogon, nogi) unikając głowy przeciwnika. Słabsze zwierzę schodzi pokonane z miejsca konfrontacji, jednak nigdy nie odnosi poważnych obrażeń – jak dotąd nie stwierdzono żadnych zranień po takich utarczkach. Rzadziej obserwuje się takie zachowania, kiedy w zbiorniku nie ma samic. Samice okazują swoją gotowość do rozrodu poprzez większą aktywność – na ogół spokojne, flegmatyczne, w obecności samca gotowego do kopulacji zaczynają okrążać go płynąc lub chodząc po dnie.
Dymorfizm płciowy
Dymorfizm płciowy u matamaty nie jest wyraźny. Samce są nieco mniejsza od samic i mają wklęsły plastron w regionie krzyżowania się tarczek femoralnych. U samicy ponadto proporcjonalnie większa jest odległość od otworu kloaki do krawędzi plastronu. Cechą charakterystyczną samców jest także często występowanie falistości (asymetrii) bruzdy środkowej plastronu. Długość ogona jest potwierdzonym kryterium szacowania płci matamaty (choć jego długość u samców jest zmienna) – w 1992 roku dowiedziono tego badaniami nad 105 muzealnymi okazami. Jednocześnie określono wklęsłość plastronu jako cechę zmienną ponieważ wystąpiła ona także u kilku samic, podczas gdy niektóre samce posiadały bardziej lub mniej płaski plastron. Mówi się także o tym, że jedne z największych samic posiadają wklęsłość plastronu w regionie środkowo abdominalnym i femoralnym (czasami są to odrębne wklęsłości w każdym z tych regionów).
Rozmnażanie
Rozród w warunkach naturalnych
Zaloty matamaty w warunkach naturalnych nie zostały opisane. Określono, że czas składania jaj u żółwi w Kolumbijskiej części Amazonki przypada na październik oraz na listopad – grudzień w Rio Putumayo. Na pojedynczy lęg przypada 12-28 jaj składanych na plażach bądź brzegach małych zatoczek. W National Zoological Park (Washington D.C.) samica złożyła 20 jaj, natomiast żółwica pochodząca z Gujany w akwaterrarium w Bronz Zoo złożyła ich aż 32. Samica w poznańskim Starym Ogrodzie Zoologicznym złożyła łącznie 23 jaja z czego 21 do wygrzebanego w podłożu dołka a dwa do wody.
Jaja tych gadów są opatrzone twardą skorupą i są niemal sferyczne. Według różnych autorów wymiary jaj wynoszą 34-40 mm, 37,2-34,3 mm (dane z populacji zasiedlających Brazylię).
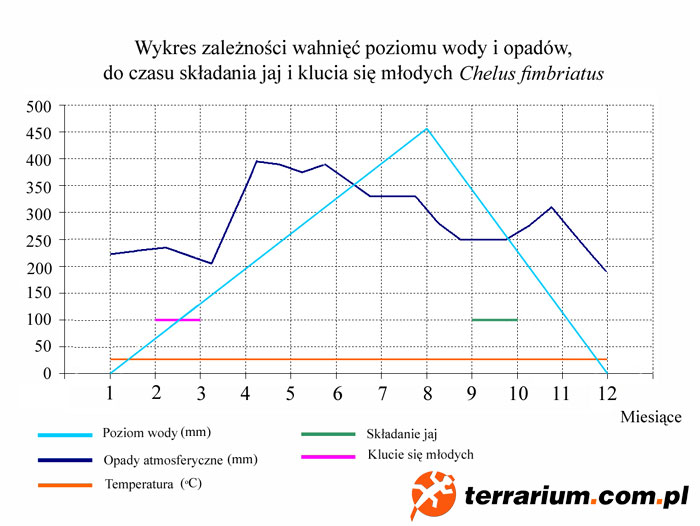
Czas inkubacji jaj może być bardzo długi i wynosi średnio 208 do nawet 9-10 miesięcy w warunkach sztucznych. W naturze inkubacja i wylęg zabierają krótszy okres czasu. Miejsca ich gniazdowania mogą być zalane w trakcie powodzi w przeciągu kilku tygodni bądź miesięcy od złożenia jaj. Możliwe, że klucie się młodych jest stymulowane przez kontakt jaja z podnoszącą się wodą podobnie jak dzieje się to w przypadku żółwi z rodzaju Carettochelys w rzekach Nowej Gwinei i północnej Australii – bez takiego kontaktu wylęg może się znacząco opóźnić. Niemniej jednak wiadome jest, że gatunek gniazduje bardzo wcześnie w porze suchej – październik lub listopad w Wenezueli co pozostawia pięć do sześciu miesięcy czasu do ponownego wezbrania wody. W Wenezueli matamaty gniazdują raczej w ilastym piasku na stromych brzegach rzek niż na łachach. W Kolumbii obserwowano matamaty wykorzystujące strome piaszczyste wybrzeża pokryte opadłymi liśćmi chętniej niż typowe piaszczyste plaże. Szukając oparcia na stromych brzegach żółwie wspinają się bokiem. Gniazda zakładane są w miejscach praktycznie niemożliwych do zlokalizowania, choć Indianie (Guahibo, Curripaco i Cuiva) bezbłędnie je znajdują. Dane te odnoszą się do rzek, niewątpliwie jednak żółwie gniazdują także w lagunach i innych miejscach.
Indianie Warrau (Warao) zasiedlają przez większą część roku obszary Plaży Almond w północno – zachodniej Gujanie. Żółwie matamata czasami pojawiały się wypełzając z morza by składać jaja na tej plaży. Takie zachowanie obserwowano we wczesnej porze suchej, po porze składania jaj żółwi morskich. Ocean w tym rejonie bywa słodkowodny zależnie od bieżących warunków oraz z uwagi na masowe odpływy z Delty Orinoko. Obserwacje te wspierają hipotezę, podług której system Essequibo zawdzięcza populację matamat raczej transportowi morskiemu z Orinoko niż przez blisko położone odpływy Amazonki i Essequibo w południowej Rupununi. Teorię tę wspiera także podobieństwo żółwi z Orinoko i Essequibo oraz ich różnic w kształcie karapaksu względem tych z Amazonki.
Gody obserwowane w warunkach sztucznych
Rodzaj specyficznych zalotów, które odbywają matamaty przed kopulacją rozpoczyna samiec, ustawiając się naprzeciwko samicy i zbliżając swój nos do jej nosa tak blisko że niemal się dotykają. Następnie samiec zaczyna powoli kołysać głową z boku na bok, zataczając przy tym lekki łuk, powtarzając to około 10 – 15 razy. Następnie zatrzymuje się chwilę w bezruchu, po czym nieznacznie podnosi jedną z przednich łap i zaczyna raptownie poruszać głowa w górę i w dół. Po około dwóch minutach samiec zaczyna kiwać głową i krążyć nerwowo po akwaterrarium. W trakcie tego chodzenia ustawia się w odpowiedniej pozycji względem samicy – prostopadle do jednego z jej boków i powtarza poprzednią procedurę. W tym czasie samica pozostaje bez reakcji. Bywa że samiec po tych zalotach podpływa nad samicę i zatrzymuje się tam na moment z szyją wyciągnięta na dół następnie odpływa bez podejmowania próby kopulacji. Przy właściwym rozwoju sytuacji samiec podpływa, bądź wspina się na samicę, następnie zagina szyję w dół i żuchwą przyciska głowę samicy. W tym czasie ogonem szuka jej kloaki. Dla stymulowania zachowań seksualnych, poleca się rozdzielać żółwie płciami na kilka miesięcy, lub też zmieniać warunki wodne i żywieniowe dla zasymulowania rytmu rocznego i sezonowości występującej w warunkach naturalnych.
Rozród w warunkach sztucznych
Do udanej reprodukcji matamaty w niewoli dochodzi bardzo rzadko, nie tylko z powodu braku seksualnej aktywności zwierząt w terrariach, ale i z powodu zamierania zarodków w lęgach przy nieprawidłowej inkubacji. Przyczyna tego leży zazwyczaj w złych warunkach bytowych – zbyt małych terrariach z źle dobranymi parametrami wody, oraz złej diecie która powoduje otłuszczanie się zwierząt. Impuls skłaniający matamaty do dobierania się w pary jest jak dotąd niepoznany, przypuszcza się, że tak jak u innych żółwi wiąże się on z rytmem rocznym, trzeba jednak jeszcze ustalić konkretnie jakie czynniki determinują u nich rozród. Średnia temperatura roczna wynosząca 26,2ºC do 27,7ºC nie wydaje się być znaczącym czynnikiem, co innego wahania poziomu wody. W zależności od szerokości rzeki i wysokości brzegów, w wyniku długotrwałych opadów różnica w poziomie wody w kolejnych sezonach może wynosić nawet 10m. Wówczas preferujące płycizny żółwie wycofują się do przybrzeżnych lagun oczekując na obniżenie poziomu wody. Prawdopodobnie więc nie głębokość jest stymulantem, a raczej sezonowo zwiększony przepływ, dlatego w akwaterrarium można to rozwiązać w następujący sposób: w okresie „zimowym” czyli kiedy w naturalnym środowisku poziom wody jest znacznie obniżony, a przepływ mniejszy, wykonuje się podmianę 15-12% wody raz w tygodniu, natomiast w okresie „letnim”, czyli kiedy w naturze występuje zwiększony przepływ, podmienia się 10% wody raz dziennie. Dodatkowym czynnikiem mogącym symulować zmienność sezonową jest dostępność pokarmu. W okresie podwyższonego poziomu wody dostępność ryb jest mniejsza, stąd racje żywieniowe żółwi hodowlanych powinny zostać obcięte do ok. 100g ryb na zwierze tygodniowo, natomiast w okresie „zimowym” kiedy poziom wody w naturze jest niski, powinny dostawać 200-300 gramowe porcje. Rezultatem takich zabiegów powinna być widoczna aktywność płciowa pod koniec okresu „letniego”, w naturze jest to przełom lutego i marca.
Lęgi w naturalnym środowisku składane są kiedy woda ponownie zaczyna opadać, co ma miejsce we wrześniu i październiku. Wylęganie się młodych przypada na początek kolejnego sezonu deszczowego, którego następstwem jest podnoszenie się poziomu wody, kalendarzowo wypada to na przełom kwietnia i maja. Składanie jaj odbywa się blisko wody, poza tym o wyborze miejsca nie wydają się decydować inne czynniki, jak nasłonecznienie czy rodzaj podłoża, którym w naturze jest najczęściej wilgotny, ubity grunt na brzegach rzek. Jaja są składane późnym wieczorem po zapadnięciu zmroku i w nocy. W terrariach jaja są zwykle zakopywane przez samice na głębokości ok. 25cm, w naturze z uwagi na mocno udeptane podłoże, które bardzo utrudnia kopanie, jaja często składane są do bardzo płytkich dołków i niemal wcale nie zakopywane.
Inkubacja jaj w warunkach hodowlanych wg różnych źródeł trwa od 180 do 207 dni, a więc dłużej niż w środowisku naturalnym Różnica ta wynika z tego, że nie zostały jeszcze zoptymalizowane warunki sztucznej inkubacji jaj – biorąc pod uwagę niedbałe zakopywanie ich przez matkę w naturalnym środowisku, podejrzewa się że to nie temperatura ma kluczowe znaczenie dla rozwijających się embrionów, a wilgotność podłoża. Substratem, który daje najlepsze jak dotąd rezultaty jest wilgotne i lekko kwaśne podłoże z mchu, utrzymywane w temperaturze 29°C ± 1°C (w naturze występują kilkustopniowe wahania między chłodniejszym porankiem i ciepłym południem). Zakwaszenie podłoża zwiększa przepuszczalność tlenu przez skorupę jaja, dzięki częściowemu rozpuszczaniu związków wapnia w niej zawartych. Ma to miejsce również w naturalnych warunkach, ponieważ wody rzek w których żyją matamaty, wody nawadniające podłoże, w którym są składane jaja, jak wspomniano wcześniej, są silnie zakwaszone.
Młode żółwiki wykluwają się w ciągu kilku minut. Po wykluciu ważą około połowy masy całego jajka czyli ok. 20g, i mierzą około 5 cm. Nos jest wygięty do tyłu, a fałdy skórne i wyrostki są schowane w szyi. Czerwony kolor spodniej części ciała jest bardzo intensywny.
Już w kilka dni po wykluciu młode zjadają małe rybki i jeżeli mają ich pod dostatkiem nie sprawiają problemu w odchowie. W naturalnym środowisku żywią się również kijankami płazów, jednak w warunkach terraryjnych raczej się tego nie stosuje, ze względu na ochronę gatunkowa rodzimych płazów i trudną dostępność tego typu komercyjnej karmówki. Przy odrobinie cierpliwości młode można łatwo przyzwyczaić do martwych, uprzednio rozmrożonych rybek, co później zasadniczo ułatwia karmienie. Najlepszym czasem na karmienie jest wczesny ranek. Jeżeli opieka jest prawidłowa, małe matamaty rosną w szybkim tempie, tak że po roku osiągają masę około 120-150 g i długość karapaksu 10-12 cm. Tak jak i dorosłe, młode można trzymać w grupie, jeżeli oczywiście zapewni im się odpowiednio duże akwaterrarium. Podobnie jak w przypadku dorosłych żółwi akwaterrarium powinno być zaopatrzone w kryjówki i różnej wysokości dno, dając żółwiom możliwość wyboru optymalnego miejsca.
Dodatkowe uwagi na temat rozmnażania matamaty
Determinacja żółwi względem składania jaj potrafi być niezwykle duża. Przykładem samica z Poznańskiego Starego Ogrodu Zoologicznego, która po złożeniu 2 jaj do wody w 2002 roku wyszła z basenu w którym była umieszczona, pokonała roślinność, wystrój z kamieni i korzeni oraz szklaną balustradę odgradzającą zbiornik od trasy dla zwiedzających. Po przejściu na drugą stronę pomieszczenia złożyła 21 jaj w samodzielnie wykopanym dołku o gruszkowatym kształcie. Niestety inkubacja nie powiodła się.
Jakkolwiek doniesienia na temat składania jaj przez żółwie matamata nie są wcale rzadkie, o tyle przypadki ich inkubacji zakończone sukcesem są niezbyt częste. Wiele jest aspektów, które mają decydujące znaczenie dla prawidłowego zapłodnienia oraz właściwej inkubacji jaj.
Z jakimi trudnościami musieli się zmierzyć hodowcy przed uzyskaniem przychówku tego gatunku, możecie przeczytać w osobnym artykule pt. Przypadek matamat Herberta Meiera (opisany wspólnie z Ingo Schaeferem).
Uwaga! Sukces reprodukcyjny matamat w warunkach sztucznych opisywany jest w literaturze rzadko. Znacznie częściej opisywano przypadki złożenia jaj. Ogólnie rzadki sukces reprodukcyjny tych zwierząt w niewoli można przypisać dwóm osobnym, podstawowym przyczynom. Po pierwsze istotne jest poznanie przyczyny wysokiego odsetka niezapłodnionych jaj, po wtóre warto wyjaśnić przypadki zamierania w pełni rozwiniętych embrionów.
Procent zapłodnionych jaj
Bez wątpliwości bardzo istotny jest stan zdrowia zwierząt przystępujących do rozrodu. Warunki utrzymania żółwi Meiera przez ostatnie lata były stale poprawiane i wyposażane zgodnie z potrzebami gadów. Do jedzenia zwierząt dodawano witaminy a świeży, żywy pokarm był podawany najczęściej jak to tylko było możliwe. Porównanie pomiędzy wzrostem masy u zwierząt utrzymywanych w hodowli i tych występujących dziko informuje o tym czy należy obciąć gadom racje żywieniowe, co oczywiście powinno być modulowane na przestrzeni roku. Ryby w zbiorniku z żółwiami służyły jako wskaźnik ilości żywności – jeśli wzrastała względem nich presja drapieżnicza ze strony żółwi, był to odpowiedni moment na ich karmienie, rosła także aktywność gadów. Równie istotna jest głębokość wody w zbiorniku. Głębokość odpowiadająca długości karapaksu największych zwierząt w zbiorniku może mieć znaczenie dla godów i udanego krycia – możliwe więc, że jest jednym z podstawowych warunków. W trakcie zalotów samiec pływa nad samicą przez kilka minut. Tę część zalotów może utrudnić lub nawet uniemożliwić przy zbyt małej głębokości zbiornika. Na głębokościach poniżej 30 cm masa samca staje się istotna w trakcie kopulacji – w takim wypadku prawidłowe krycie staje się trudne albo niewykonalne. Ważny jest także sam rozmiar zbiornika dla żółwi. Matamaty często bywają trzymane w zbiornikach daleko dla nich za małych. Naturalne zachowania rozrodcze tych zwierząt w takim wypadku są utrudnione lub niemożliwe.
Procent wylęgających się młodych
Wielu autorów/hodowców obserwowało, że embriony zamierają w jaju i to na późnym etapie rozwoju, będąc niemal całkowicie wykształcone. Wielu z nich twierdzi także, że jaja składane są przez okres kilku dni lub nawet tygodni lub że złożenie ich ma miejsce w wodzie. Oczywistym powodem wydaje się tu być miejsce udostępnione żółwiom dla celów gniazdowych – może ono zwyczajnie nie spełniać ich wymagań i preferencji. To z kolej prowadzi do opóźnienia skutkującego nadmiernym zwapnieniem jaj, co w konsekwencji utrudnia dyfuzję tlenu przez skorupę jaja. Można tego uniknąć jedynie po przez stwarzanie żółwiom prawidłowych warunków lęgowych.
Nawet jeżeli w zbiorniku jest już odpowiednie miejsce, które zostało zaakceptowane przez matamaty, procent klujących się młodych może nie być satysfakcjonujący. Zjawisko to możne mieć różne przyczyny. Prostym czynnikiem przeciwdziałającym temu zjawisku może być utrzymanie właściwej żywotności (witalności) rodziców. Niestety różnorodność ryb, którymi można karmić matamaty jest ograniczona. W większości przypadków należą one do ryb karpiowatych w szerszym sensie. Ponadto są one pozyskiwane (tak jak w przypadku Meiera) z farm o zintensyfikowanych metodach chowu i przechowywane w zamrażarkach. Problem ten rozwiązano podając w ostatnich latach mrożone okonie pochodzące ze stawu o ekstensywnym sposobie użytkowania. Regularne porcje witamin rekompensują wszelkie nieprawidłowości żywieniowe mogące wystąpić w warunkach sztucznych. Głównym powodem niskiego procenta wyklutych żółwi może być jednak metoda inkubacji. Doświadczenia hodowców potwierdzają, że inkubacja może przebiec prawidłowo o ile jaja zostaną ułożone na lub w wilgotnym mchu. Brak lub zły typ substratu może utrudnić erozję wapnia w skorupach jaj, co w konsekwencji może być przyczyną trudności w wyjściu z jaj młodych żółwi. Wylęg może być ponadto utrudniony przez ograniczoną dyfuzję tlenu przez skorupy nienaruszonych (mocno zwapniałych) jaj. W każdym wypadku obserwacje potwierdzają, że lekko kwaśne środowisko inkubacji jaj jest korzystne dla prawidłowego ich wylęgu. Różni autorzy podają odmienne czasy inkubacji jaj. Zakres ten mieści się w przedziale 168 – 300 dni. Tak duże rozbieżności w czasie inkubacji są prawdopodobnie spowodowane różnym stopniem zawilgocenia substratu inkubacyjnego, nie natomiast samą temperaturą inkubacji. Niestety informacje na temat wilgotności panującej w substracie także są rozbieżne – stosowano zarówno relatywnie suchy jak i zdecydowanie wilgotny. Otwartym pytaniem pozostaje więc w jaki sposób wilgotność substratu inkubacyjnego może przyspieszać rozwój zarodków. Może to być mechanizm zawiadujący procesem wylęgu młodych żółwi. Udowodniono jednak że przesuszenie substratu może być czynnikiem przyspieszającym czas inkubacji jaj żółwi z gatunku Emys orbicularis i Emudura subglobosa.
Wnioski. Jeżeli Chelus fimbriata utrzymywany jest w odpowiednich warunkach, egzystuje w zbiorniku dostosowanym wielkością do jego potrzeb, złożenie jaj może zostać zrealizowane relatywnie niedługo po okresie aklimatyzacji. Procent zapłodnionych jaj jak i wykluwających się młodych nadal nie są satysfakcjonujące i pozostawiają wiele pytań. Sukces wylęgu jest wspomagany przez nieznaczne zakwaszony substrat inkubacyjny jak, na przykład wilgotny mech, którym jaja powinny być całkowicie pokryte. Wilgotność powinna wzrosnąć przy końcu inkubacji co może wpływać korzystnie na przebieg wylęgu. Odchów młodych nie nastręcza problemów, poza przyzwyczajeniem ich do przyjmowania martwego pokarmu.
Młode
Masa młodych urodzonych w niewoli (dane z 1968 roku) to ok. 15 g, przy czym w przeciągu kolejnych 2 miesięcy wzrasta ona do ok. 30 g. Według nowszych danych (1996) młode mają ok. 49 mm CL i ok. 34 cm szerokości i masę 15 – 19 g. Sześciomiesięczne młode mają 60 g i około 80 mm CL.
Młode żółwie matamata w warunkach sztucznych szybko rosną. W pierwszym miesiącu życia przybierają ok. 10 g miesięcznie (począwszy od masy ok. <20g). Ich kolor w tym okresie także ulega zmianom – często w pierwszych miesiącach życia zyskują na kontraście jednak u starszych jaskrawe ubarwienie nie jest widoczne poza spodem szyi w ograniczonym stopniu. W akwaterrarium wybierają swoje ulubione miejsca, najczęściej te umożliwiające schowanie znacznej części pancerza pod kamieniem lub korzeniem tak by móc wystawić nos nad wodę bez przemieszczania się. Czasem kopią w podłożu. Są stosunkowo aktywne, zwłaszcza w godzinach późno-popołudniowych i nocnych. Gdy są głodne potrafią polować aktywnie. Należy pamiętać, że mamy do czynienia z gatunkiem rzecznym. Nie ma potrzeby utrzymywania wysokich temperatur. Młode żółwie dobrze rozwijają się w temperaturze ok. 25°C.
Status populacji w środowisku naturalnym
Nie ma danych ilościowych na temat populacji tego żółwia. W ogólnym zakresie należy stwierdzić, że gatunek jest raczej rzadko widywany lub napotykany przypadkiem i odnosi się to raczej do pojedynczych egzemplarzy. Może to być jednak wynikiem raczej jego skrytego trybu życia niż rzadkości bądź niskiego zagęszczenia. Napotkano nienaturalne zagęszczenie tego gatunku w sieci wód sezonowo płynących w porze suchej w Rio Tinaco, dopływie Orinoko w Rancho la Trinidad, Estado Cojedes w Wenezueli. W tej lokalizacji złapano 11 dużych okazów za pierwszym podejściem i w przeciągu 24 godzin 24 dorosłe żółwie po tej samej stronie cieku wodnego.
Komercyjny wywóz matamat sugeruje że kolekcjonerzy odkryli korzyści płynące z ich licznego importu. Na przykład, choć gatunek jest określany jako rzadki w Gujanie i jest zaledwie kilka okazów muzealnych z tej lokalizacji w 1992 roku wywieziono stamtąd ok. 60 dorosłych żółwi do USA, a mniej więcej dekadę wcześniej kilkaset osobników młodocianych zostało nielegalnie importowanych do Kalifornii, gdzie zostały skonfiskowane, a następnie trafiły do Zoo w Los Angeles i zostały rozdysponowane do innych ogrodów zoologicznych i kolekcji. W Kolumbii w sieci rybackie dostaje się wiele młodych żółwi, zwłaszcza w sąsiedztwie Puerto Inírida w Gujanie i okolicy Caño Cabuyaro, dopływu Górnej Meta (Meta/Casanare).
Wrogowie naturalni
Matamata posiada wrogów na wolności. W Karanambo i Gujanie na rzece Rupununi zaobserwowano dorosłe matamaty regularnie wyciągane na brzeg i zabijane przez wydry rzeczne (ariranie) [pancerz 38,8 cm długości dojrzałego samca matamaty z Karanambo nosił znaki tego, że zwierze padło łupem wydry rzecznej]. W Estado Cojedes (Wenezuela) wszystkie prócz 5 z 24 żółwi złapanych w porze suchej w okolicy llanos nosiły mniejsze lub większe blizny i okaleczenia: brakujące palce we wszystkich przypadkach, czasem brak całej kończyny, odgryzioną trąbkę (nos) czy płaty skóry okolic głowy i szyi. Wiele ryb drapieżnych (łącznie z licznie występującą piranią Serrasalmo sp.) dzieli przestrzeń życiową z żółwiem matamata i to właśnie one mogą być odpowiedzialne za tego typu okaleczenia.
Zagrożenia w środowisku naturalnym
Matamata będąc gatunkiem kryptycznym i szeroko rozprzestrzenionym o małym znaczeniu dla człowieka (z uwagi na walory konsumpcyjne) nie wydaje się być bezpośrednio zagrożony. W rejonach jego występowania obecne są inne, alternatywne gatunki żółwi (zwłaszcza rodzaje Podocnemis i Chelonoidis – oba cenione z uwagi na aspekty konsumpcyjne). Lokalna ludność raczej nie jada matamaty z uwagi na jego zapach i wygląd – wolą żółto-cętkowanego żółwia rzecznego Podocnemis unifilis bądź południowo amerykańskiego żółwia rzecznego Podocnemis expansa. Ostatni z tych gatunków w konsekwencji niemal zniknął z rzek llanos. Świeżo złapany matamata wydziela stosunkowo brzydki zapach, jest na tyle osobliwy w wyglądzie i ma nietypowo ulokowane mięso nadające się do spożycia (głównie w obszarze szyi, kończyny są małe i słabe) że większość ludności plemiennej i wiejskiej w Wenezueli odrzuca go jako „la fea” (brzydkiego) i zjada inne gatunki bardziej „normalnie” wyglądających żółwi. Jednak we wcześniejszym okresie oraz innych miejscach nie to mogło mieć główne znaczenie. Niektórzy autorzy donoszą, że matamata był intensywnie poszukiwany z uwagi na swoje mięso w okolicach Ile de Cayenne że stał się zagrożony i nie można go było znaleźć w znacznej ilości w odległości bliższej niż 25 mil na południe od miasta.
Jako żywa ciekawostka matamata jest chętnie widywany przez hobbystów w Europie i Stanach Zjednoczonych. Zainteresowanie koncentruje się głównie na osobnikach młodocianych. Eksporty rzędu 60 dorosłych dużych egzemplarzy z Gujany na Florydę w 1992 roku opatrzone były wysokimi cenami za sztukę (ok. $1400 US), lecz sprzedaż nie osiągała wysokiego pułapu a rynek na te drogie i duże żółwie szybko się nasycił.
Podjęte środki ochrony
Nie podjęto działań mających na celu ochronę tego gatunku żółwia, choć jest on teoretycznie chroniony po przez kompleksową ochronę dzikiej przyrody (prawo i zakazy na eksport w Kolumbii, Wenezueli i Brazylii). W kilku przypadkach populacje matamat mogą być przedmiotem ochrony z racji występowania na terenie Parku Narodowego lub innego obszaru ochrony (jak Xingu w Brazylii, Manu w Peru, Iwokrama w Gujanie i Canaima w Wenezueli).
Żółw nie znajduje się na Czerwonej Liście IUCN, ani na załączniku Konwencji Waszyngtońskiej CITES. Zawarty jest w kategorii NT (ang. Near Threatened – bliski zagrożenia) na Kolumbijskiej Czerwonej Liście (Castaño-Mora 2002). Nie ma go na Czerwonej Liście Wenezueli (Rodriguez and Rojas-Suarez 1999). Matamata chroniony jest francuskim prawem w Gujanie Francuskiej, którego egzekwowanie jest jednak nieadekwatne.
Proponowane środki ochrony
Nie jest wymagane podjęcie natychmiastowych działań w kwestii ochrony tego gatunku, choć niektóre czynniki, zwłaszcza międzynarodowy handel żywymi zwierzętami oraz degradacja środowiska naturalnego powinny być monitorowane.
Opracowanie i źródła informacji
Agnieszka Graclik
Instytut Zoologii, Zakład Zoologii
Przemysław Szwajkowski
Zakład Agroturystyki
Wydział Hodowli i Biologii Zwierząt
Uniwersytet Przyrodniczy w Poznaniu
na podstawie doświadczeń własnych i literatury:
„Aditional Remarks on the Breeding of Chelus fimbriatus – H. Meier, I. Schaefer. 2003. Radiata 12 (4).
„Bizarre Nature The Mata Mata” – C.L. Barrio-Amorós. Reptilia vol.
„ Chelus fimbriatus (Shneider 1783) – Matamata Turtle” – P.C.H. Pritchard. 2008. Conservation Biology of Freshwater Turtles and Tortoises. August.
„Experiences and observations of Chelus fimbriatus, the maintenance in terrarium and successful reproduction” – H. Meier, I. Schaefer. 2003. AG Schildkröten in Deutsche Gesellschaft fur Herpetologie und Terrarienkunde e. V. (DGHT)
„Matamata Chelus fimbriatus by Zig Leszczynski” – Z. Leszczynski. 2006. Reptiles, January.
„Turtles of Venezuela (Conributions to herpetology)” – P.C.H. Pritchard, Trebbau. 1984. Society of the Study of Amphibians and Reptiles.
„Turtles of the World” – R.W. Barbour, C.H. Ernst. 1992. Smithsonian Institution Press.
„What’s the Mata?” – D. Fogel. 2006. Reptiles magazine, No. 5. Published 4 January.
![]() Copyright by Agnieszka Graclik & Przemysław Szwajkowski 2010
Copyright by Agnieszka Graclik & Przemysław Szwajkowski 2010
Wszystkie prawa zastrzeżone. Zgodnie z prawem o ochronie praw autorskich do własności intelektualnej [Dz.U.1994r.Nr24poz.83; Dz.U.2000r.Nr80poz.904] zabrania się, bez zgody obu autorów, modyfikowania, kopiowania i rozpowszechniania całości bądź też fragmentów niniejszej pracy. Zastrzeżono nienaruszalność treści i formy artykułu.
Liczba wyświetleń: 14505


















